

A Resveratrol and Polyphenol Preparation Suppresses
Oxidative and Inflammatory Stress Response
to a High-Fat, High-Carbohydrate MealThis section is compiled by Frank M. Painter, D.C.
Send all comments or additions to: Frankp@chiro.org


FROM: J Clin Endocrinol Metab. 2011 (May); 96 (5): 1409–1414 ~ FULL TEXT
Husam Ghanim, Chang Ling Sia, Kelly Korzeniewski, Teekam Lohano,
Sanaa Abuaysheh, Anuritha Marumganti, Ajay Chaudhuri and Paresh Dandona
Division of Endocrinology,
Diabetes, and Metabolism,
State University of New York at Buffalo,
and Kaleida Health,
Buffalo, New York 14209.
Did you see the Oscar-nominated movie Superzize Me? If so, you will recall how Morgan Spurlock’s health declined during the month he lived on fast food. The saturated and trans-fats, preservatives, and other food additives contained within fast food meals depress immune function, cause arterial constriction, depose plaque in the arteries, and increases cortisol production, leading to stress reactions and accelerated aging. That’s why Morgan’s health declined. And you saw just how fast that happened.
In this randomized, placebo-controlled clinical study, performed by scientists at State University of New York at Buffalo, participants were fed a 910–calorie, fast food breakfast, including sausage, cheese and fried potatoes (a Burger King-style breakfast). Half the group received a resveratrol and polyphenol supplement. Everyone’s blood was tested for the activity levels of a key genetic molecule, called Nrf2 (pronounced Nerf 2), which is the coordinator of antioxidant protection within every cell in your body. The Nrf2 defense system mobilizes antioxidant enzymes, but it also choreographs immune response, and enhances cellular detoxification. So disruption of this system spells disaster for the body.
Nrf2 activity was immediately depressed in the control group, while the supplemented group demonstrated dramatic increases in Nrf2 activity. And YES, this is very important because it helps to explain how poor dietary choices dramatically affect our health. They do so by suppressing the body’s natural defense mechanisms.
Let’s review the amazing characteristics of the Nrf2 defense system:. Phase II detox enables the body to excrete xenobiotics, or non-human compounds, like drugs and other toxic chemicals, that Phase I liver detox cannot eliminate. Depression of Nrf2 activity leaves the body less capable of removing these toxins. [1]Nrf2 manages Phase II detoxification within the liver
Disruption of the Nrf2 gene leads to severe allergen-driven airway inflammation that contributes to the development of asthma. [2]
Nrf2 has also just been found to inhibit the development of emphysema by researchers at Johns Hopkins’s School of Public Health. [3] Four years ago, this same group at Johns Hopkins demonstrated that Nrf2 protected against developing pulmonary fibrosis, a frequently fatal lung disease. [4–5]Because oxidative stress is associated with the onset of so many different disease processes, the Nrf2 antioxidant system has recently been found to be associated with reduced risk for developing Parkinson's [6] and Huntington's disease [7], lung cancer [8], and other inflammatory and autoimmune diseases. [9] It’s also been shown to provide protection against a variety of pathologies in numerous organs, including the liver, intestine, lung, skin, and nervous system. [10]
This clinical research study demonstrated that a unique nutritional supplement, containing resveratrol and muscadine grape polyphenols can blunt the biological stress response that occurs after consuming a high-fat, high-carbohydrate meal (HFHC). Researchers gave the participants either the resveratrol/polyphenol combination or a placebo just before consuming a 930–kcal HFHC meal, and then then observed the metabolic responses in their bloodstream over a 5–hour period. It is well-known that HCHC meals induce significant oxidative and inflammatory stress.
Results showed significant blunting of the typical oxidative and biological stress response, as well as significant activation of Nrf-2, a key regulator of our antioxidant defense system, in the group consuming the supplemented polyphenols.
This research nicely complements another mitochondrial mass study showing increased cellular energy production with supplemented resveratrol/polyphenols compared to resveratrol alone, and another laboratory study that showed this combination supplement was more powerful than resveratrol alone at inhibiting AGE protein formation.
REFERENCES:
Disruption of Nrf2 enhances susceptibility to severe airway inflammation and asthma in mice
J Exp Med. 2005 Jul 4;202(1):47-59Activating the Lung's Antioxidant Defense By Targeting Nrf2 Inhibits the Development of Emphysema
Medical News Today 2008 (Dec 29)
Johns Hopkins Bloomberg School of Public HealthThe transcription factor NRF2 protects against pulmonary fibrosis
FASEB Journal 2004 (Aug); 18 (11): 1258-1260Nrf2-mediated neuroprotection in the MPTP mouse model of Parkinson's disease:
Critical role for the astrocyte
Proc Natl Acad Sci U S A. 2009 (Feb 24);106(8):2933-8Mutant huntingtin activates Nrf2-responsive genes and impairs dopamine synthesis in a
PC12 model of Huntington's disease
BMC Mol Biol 2008 (Oct 9); 9: 84Dysfunctional KEAP1-NRF2 interaction in non-small-cell lung cancer
PLoS Med 2006 (Oct); 3 (10): e420Protective Roles of Nrf2 in Disease including Oral Disease
J Oral Biosciences 2005; 47 (2) 126-134Emerging Role of Nrf2 in Protecting Against Hepatic and Gastrointestinal Disease
Toxicol Pathol 2007; 35 (4): 459-73
Background: High-fat, high-carbohydrate (HFHC) meals are known to induce oxidative and inflammatory stress, an increase in plasma endotoxin concentrations, and an increase in the expression of suppressor of cytokine signaling-3 (SOCS-3).
Hypothesis: The intake of a nutritional supplement containing resveratrol and muscadine grape polyphenols reduces HFHC meal-induced oxidative and inflammatory stress and stimulates the activity of the antioxidant transcription factor, NF-E2-related factor-2 (Nrf-2), and its downstream targets.
Methods: Ten normal, healthy subjects were given a 930–kcal HFHC meal either with placebo or with the supplement. Indices of oxidative stress, inflammation, Nrf-2 binding activity, the concentrations of endotoxin (lipopolysaccharide) and lipoprotein binding protein (LBP), and the expression of toll-like receptor 4 (TLR-4), CD14, IL-1ß, TNFa, SOCS-3, Keap-1, NAD(P)H:quinone oxidoreductase-1 (NQO-1), and GST-P1 were measured.
Results: The intake of the supplement suppressed the meal-induced elevations of plasma endotoxin and LBP concentrations, the expression of p47(phox), TLR-4, CD14, SOCS-3, IL-1ß, and Keap-1, while enhancing Nrf-2 binding activity and the expression of NQO-1 and GST-P1 genes.
Conclusion: A supplement containing resveratrol and muscadine polyphenols suppresses the increase in oxidative stress, lipopolysaccharide and LBP concentrations, and expression of TLR-4, CD14, IL-1ß and SOCS-3 in mononuclear cells after an HFHC meal. It also stimulates specific Nrf-2 activity and induces the expression of the related antioxidant genes, NQO-1 and GST-P1. These results demonstrate the acute antioxidant and antiinflammatory effects of resveratrol and polyphenolic compounds in humans in the postprandial state.
From the Full-Text Article:
Introduction:
The intake of a high-fat, high-carbohydrate (HFHC) meal induces oxidative and inflammatory stress as reflected by increased reactive oxygen species (ROS) generation, increased expression of the subunits of reduced nicotinamide adenine dinucleotide phosphate (NADPH) oxidase, increased nuclear factor-ΚB binding in mononuclear cells (MNC), and increased expression and plasma concentrations of proinflammatory cytokines. [1] Such meals also elicit an increase in plasma concentrations of endotoxin [lipopolysaccharide (LPS)] and lipoprotein binding protein (LBP) and the expression of its receptor, toll-like receptor-4 (TLR-4). [2] Our recent work has shown that HFHC meals also evoke an increase in the expression of the suppressor of cytokine signaling-3 (SOCS-3), a key protein responsible for interference with insulin and leptin signal transduction. [3–5]
One potent antioxidant mechanism triggered by oxidative stress is that based on the induction of the transcription factor NF-E2-related factor-2 (Nrf-2), which activates the transcription of a series of genes that have an antioxidant response element (ARE) in their promoters. [6] These genes include NAD(P)H:quinone oxidoreductase-1 (NQO-1), glutathione S-transferase-1P (GST-1P), superoxide dismutase (SOD)-2, thioredoxin, and glutamate cysteine ligase-2 (GCL-2). All of these proteins exert potent antioxidant effects by increasing the clearance or neutralization of ROS. The transcriptional activity of Nrf-2 is regulated by Keap-1, a cytosolic protein that binds to Nrf-2 and thus prevents its translocation into the nucleus. An increase in Keap-1 expression is therefore inhibitory to Nrf-2 activity, whereas a decrease enhances Nrf-2 activity. Nrf-2 has recently been shown to protect vascular cells from oxidative stress. [7, 8] There are hitherto no data on the action or the modulation of Nrf-2 in humans in vivo.
We have recently shown that an extract of Polygonum cuspidatum containing resveratrol suppresses ROS generation, p47phox and SOCS-3 expression, and nuclear factor-ΚB binding when given to normal, healthy subjects (at 40 mg/d) over a period of 6 wk. [9] In view of the above, we have now hypothesized that the intake of a preparation containing resveratrol and polyphenols from muscadine grapes reduces or prevents HFHC meal-induced oxidative and inflammatory stress and the induction of SOCS-3. In addition, it induces Nrf-2 activity and the transcription of its target antioxidant genes.
Subjects and Methods:
Subjects
A group of 10 (six females) normal-weight, healthy, nonsmoking subjects (age, 37 ± 4 yr; body mass index, 22.6 ± 0.5 kg/m2; blood pressure, 117 ± 4 and 74 ± 3 mm Hg; fasting glucose concentration, 86 ± 9 mg/dl; fasting insulin, 6.5 ± 9 µU/ml; triglycerides, 68 ± 9 mg/dl; total cholesterol, 138 ± 11 mg/dl) were recruited for this crossover and placebo-controlled study. After an overnight fast, subjects were given, on two separate visits 1 wk apart, a 930–calorie HFHC meal [2] with either a single dose of a nutraceutical supplement or a placebo 10 min before the meal. The supplement consisted of 100 mg of resveratrol from P. cuspidatum extract plus 75 mg of total polyphenols from a muscadine grape extract (Shaklee Corporation, Pleasanton, CA). Blood samples were collected at baseline and at 1, 3, and 5 h after meal intake. The experimental protocol was approved by the Human Research Committee of the State University of New York at Buffalo, and each subject signed an informed consent.
MNC isolation, Western blotting, and RT-PCR
MNC and polymorph nuclear cells were isolated [1], and protein and mRNA expression [2, 10] was determined as previously described. Antibodies against TLR-4, SOCS-3, Keap-1 (Abcam Inc., Cambridge, MA), CD14, and actin (Santa Cruz Biotechnology, Santa Cruz, CA) were used. Expression of Nrf-2, NQO1, GST, SOD, and IL-1ß mRNA was measured using specific primers (Invitrogen, Carlsbad, CA).
Nrf-2 DNA binding activity
Nuclear Nrf-2 DNA binding activity was measured by EMSA as described previously. [1, 11] Oligonucleotide corresponding to the Nrf-2 binding sites on NQO1 promoter [12] (sense, GTCAGTGTCACTGAGTCGTCTTAGA; and antisense, CAGTCACAGTGACTCAGCAGAATCT) was used. The specificity of the band was confirmed by supershifting the band with specific antibody against Nrf-2 (Santa Cruz Biotechnology) and by competition with cold oligonucleotides.
Plasma endotoxin and LBP concentrations were measured as previously described. [10]
Statistical analysis
Statistical analysis was conducted using SigmaStat software version 3.1 (SPSS Inc., Chicago, IL). Data are expressed as mean ± se. Percentage change from baseline was calculated, and statistical analysis for change from baseline was carried out using one-way repeated measures ANOVA (RMANOVA) followed by Holm-Sidak post hoc test. Two-factor RMANOVA analysis followed by Tukey's post hoc test was used for all multiple comparisons between groups.
Results:
Supplement effect on meal-induced oxidative stress
After consumption of the meal, MNC protein levels of p47phox (NADPH subunit) increased by 148 ± 38% over baseline (P < 0.05) in the placebo group, whereas no significant changes were observed in the supplement group (Fig. 1, A and B). The DNA binding activity of the antioxidant transcription factor Nrf-2 was significantly increased by 150 ± 39% over baseline (P < 0.05) at 3 h after meal and supplement intake, whereas meal consumption in the placebo group resulted in a significant reduction in Nrf-2 binding activity at 5 h (Fig. 1, C and D). This was associated with a significant reduction by 48 ± 6% (P < 0.05) in the supplement group and a significant increase by 66 ± 10% (P < 0.05) in the supplement group in Nrf-2 inhibitor, Keap-1, protein levels in MNC (Fig. 1, A and E). Additionally, significant increases (P < 0.05) in the mRNA expression of Nrf-2 target genes, NQO-1 and GST-P1 (data not shown), and in NQO-1 protein levels (Fig. 1, A and F) were observed after the meal plus supplement, whereas there was no change in the expression of NQO-1 or GST-P1 after the meal plus placebo. There was no change in the expression of other Nrf-2 target genes including SOD-2, thioredoxin, or GCL in either group.
Supplement effect on meal-induced inflammation
There was a significant, steady increase in plasma concentrations of LPS in the placebo group from 0.23 ± 0.02 endotoxin units (EU)/ml to 0.29 ± 0.03, 0.33 ± 0.04, and 0.36 ± 0.03 EU/ml at 1, 3, and 5 h, respectively, after the meal (60 ± 16% over baseline at 5 h; P < 0.05). In contrast, plasma LPS concentrations decreased significantly by 28 ± 7% below baseline at 1 h (from 0.30 ± 0.04 EU/ml to 0.22 ± 0.04, 0.33 ± 0.04, and 0.34 ± 0.05 EU/ml at 1, 3, and 5 h, respectively) in the supplement group. This was associated with an increase in plasma levels of the LBP by 38 ± 16% over the baseline at 3 h (from 10.78 ± 1.2 to 15.6 ± 2.4 µg/ml; P < 0.05; Fig. 2A) in the placebo group; this increase was prevented by the intake of the nutraceutical supplement. The intake of the HFHC meal in the placebo group induced a significant increase in mRNA expression of IL-1ß by 91 ± 28% (from 0.34 ± 0.08 to 0.61 ± 0.14 relative units compared with control sample; P < 0.05), whereas this increase was significantly reduced in the supplement group to 29 ± 11% over the baseline (from 0.38 ± 0.1 to 0.47 ± 0.11 relative units compared with control sample; P < 0.05) (Fig. 2B).
Expression of SOCS-3, TLR-4, and CD14 proteins
As shown in Fig. 2, C and D, SOCS-3 protein levels in MNC increased by 39 ± 13% (P < 0.05) over baseline in the placebo group, whereas no significant changes in SOCS-3 protein levels were measured in the supplement group. Similarly, there was a significant increase in the expression of CD14 and TLR-4 protein by 67 ± 19 and 40 ± 15%, respectively (P < 0.05), over baseline after the HFHC meal, which was prevented by the concomitant intake of the supplement but not the placebo (Fig. 2 C, E and F).
Figure 1. Change from baseline
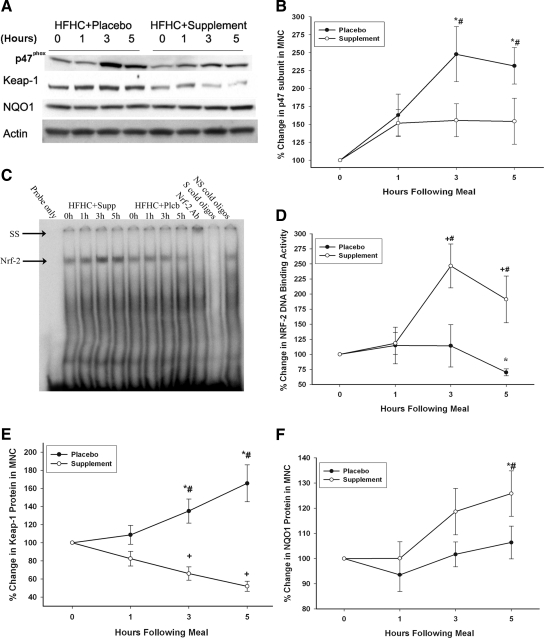
Figure 2. Change in LBP
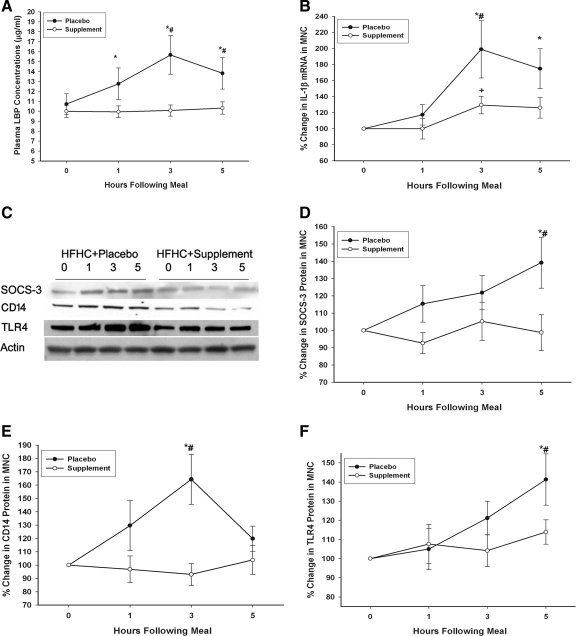
], the consumption of the HFHC meal with the placebo produced an increase in the expression of p47phox, an essential subunit of NADPH oxidase that generates the O2 radical and mediates oxidative stress. The HFHC meal also induced an increase in LPS and LBP concentration and the expression of TLR-4, CD14, and SOCS-3, all of which were either reduced or totally prevented by the supplement. The prevention of an increase in LPS is intriguing but is consistent with the action of orange juice, which we described recently [9]. It is possible that resveratrol, the muscadine polyphenols, and orange juice may exert a direct protective antiinflammatory effect on the intestinal epithelium, including any change in intestinal permeability. This issue requires further investigation.
Discussion:
As demonstrated previously [1, 2
The induction of CD14 and TLR-4 along with an increase in LPS and LBP by the meal has implications for the pathogenesis of both atherogenesis and insulin resistance. The deletion of TLR-4 protects against both insulin resistance in mice fed a high-fat diet [13] and atherosclerosis in mice with apolipoprotein E deletion. In this context, there is a significant expression of TLR-4 in cells of the atherosclerotic plaque [14].
Our other major novel observation is that there was a significant increase in the intranuclear binding activity of Nrf-2 at 3 and 5 h after consumption of the HFHC meal with the supplement. In contrast, Nrf-2 binding activity actually decreased (at 5 h) in the placebo group. Nrf-2 is an important transcription factor that mediates protective responses to oxidative stress by binding to the ARE of several genes, including those responsible for glutathione synthesis and phase II drug metabolism [6]. Consistent with this, the expression of Keap-1, the protein to which Nrf-2 is bound in the cytosol and which also causes its ubiquitination and proteasomal degradation, was reduced markedly in the group receiving the supplement, whereas it increased significantly in the placebo group. The decline in Keap-1 levels would potentially stabilize Nrf-2, allow its translocation into the nucleus, and facilitate the initiation of transcription of multiple protective antioxidant genes including NQO-1 and GST-1P, two major enzymes with antioxidant activity. Nrf-2 has also been shown to reverse biochemical dysfunction in endothelial cells induced by high glucose concentrations [8]. Nrf-2 expression and activity have recently been shown to be diminished with aging [15].
The prevention of the increase in SOCS-3, a protein that interferes with insulin and leptin signal transductions, after the HFHC meal in the supplement group suggests that the supplement may have a role in preventing insulin and leptin resistance in long-term studies [2]. The expression of SOCS-3 has been shown to be elevated in obese humans in whom it is inversely related to insulin receptor phosphorylation [16]. The prevention of the induction of IL-1ß by the intake of the supplement is also potentially insulin sensitizing because IL-1ß in turn induces the expression of SOCS-3.
In conclusion, the intake of a resveratrol and polyphenol-based nutritional supplement before an HFHC meal significantly reduced multiple indices of oxidative and inflammatory stress: p47phox, IL-1ß, CD14, and TLR-4 expression and LPS and LBP concentrations. The intake of the supplement also induced the key antioxidant transcription factor, Nrf-2, and its gene targets, NQO-1 and GST-1P, while suppressing Keap-1, the antagonist of Nrf-2.
Acknowledgments
The authors thank Elizabeth Szudzik, Barbara Hall, and Lisa Martin for their invaluable help with this study.
The study was supported by a grant from Shaklee Corporation.
Disclosure Summary:
P.D. is also supported by National Institutes of Health Grants R01DK069805-02 and R01DK075877-01-A2; and American Diabetes Association grant 08-CR-13. P.D. has received research support and lecture honoraria from GSK and Sanofi-Aventis. A.C. has received lecture honoraria from Eli Lilly. H.G., C.S., K.K., T.K., S.A., and A.M. have nothing to disclose.
References :
Aljada, A, , Mohanty, P, , Ghanim, H, , Abdo, T, , Tripathy, D, , Chaudhuri, A, 2004
Increase in intranuclear nuclear factor ΚB and decrease in inhibitor ΚB in mononuclear
cells after a mixed meal: evidence for a proinflammatory effect.
Am J Clin Nutr 79:682–690Ghanim, H, , Abuaysheh, S, , Sia, CL, , Korzeniewski, K, , Chaudhuri, A, 2009
Increase in plasma endotoxin concentrations and the expression of Toll-like receptors
and suppressor of cytokine signaling-3 in mononuclear cells after a high-fat,
high-carbohydrate meal: implications for insulin resistance.
Diabetes Care 32:2281–2287Emanuelli, B, , Peraldi, P, , Filloux, C, , Chavey, C, , Freidinger, K, , Hilton, DJ, 2001
SOCS-3 inhibits insulin signaling and is up-regulated in response to tumor necrosis
factor-a in the adipose tissue of obese mice.
J Biol Chem 276:47944–47949Rui, L, , Yuan, M, , Frantz, D, , Shoelson, S, , White, MF, 2002
SOCS-1 and SOCS-3 block insulin signaling by ubiquitin-mediated degradation of
IRS1 and IRS2.
J Biol Chem 277:42394–42398Bjřrbaek, C, , El-Haschimi, K, , Frantz, JD, , Flier, JS, 1999
The role of SOCS-3 in leptin signaling and leptin resistance.
J Biol Chem 274:30059–30065Kang, KW, , Lee, SJ, , Kim, SG, 2005
Molecular mechanism of nrf2 activation by oxidative stress.
Antioxid Redox Signal 7:1664–1673Kim, SH, , Kim, SH, , Choi, M, , Lee, Y, , Kim, YO, , Ahn, DS, 2010
Natural therapeutic magnesium lithospermate B potently protects the endothelium
from hyperglycaemia-induced dysfunction.
Cardiovasc Res 87:713–722Xue, M, , Qian, Q, , Adaikalakoteswari, A, , Rabbani, N, 2008
Activation of NF-E2-related factor-2 reverses biochemical dysfunction of endothelial
cells induced by hyperglycemia linked to vascular disease.
Diabetes 57:2809–2817Ghanim, H, , Sia, CL, , Abuaysheh, S, , Korzeniewski, K, , Patnaik, P, 2010
An antiinflammatory and reactive oxygen species suppressive effects of an extract
of Polygonum cuspidatum containing resveratrol.
J Clin Endocrinol Metab 95:E1–E8Ghanim, H, , Sia, CL, , Upadhyay, M, , Korzeniewski, K, , Viswanathan, P, 2010
Orange juice neutralizes the proinflammatory effect of a high-fat, high-carbohydrate
meal and prevents endotoxin increase and Toll-like receptor expression.
Am J Clin Nutr 91:940–949Andrews, NC, , Faller, DV, 1991
A rapid micropreparation technique for extraction of DNA-binding proteins
from limiting numbers of mammalian cells.
Nucleic Acids Res 19:2499Tirumalai, R, , Rajesh Kumar, T, , Mai, KH, , Biswal, S, 2002
Acrolein causes transcriptional induction of phase II genes by activation of Nrf2
in human lung type II epithelial (A549) cells.
Toxicol Lett 132:27–36Shi, H, , Kokoeva, MV, , Inouye, K, , Tzameli, I, , Yin, H, , Flier, JS, 2006
TLR4 links innate immunity and fatty acid-induced insulin resistance.
J Clin Invest 116:3015–3025Xu, XH, , Shah, PK, , Faure, E, , Equils, O, , Thomas, L, , Fishbein, MC, 2001
Toll-like receptor-4 is expressed by macrophages in murine and human lipid-rich
atherosclerotic plaques and upregulated by oxidized LDL.
Circulation 104:3103–3108Suh, JH, , Shenvi, SV, , Dixon, BM, , Liu, H, , Jaiswal, AK, , Liu, RM, , Hagen, TM, 2004
Decline in transcriptional activity of Nrf2 causes age-related loss of glutathione synthesis,
which is reversible with lipoic acid.
Proc Natl Acad Sci USA 101:3381–3386Ghanim, H, , Aljada, A, , Daoud, N, , Deopurkar, R, , Chaudhuri, A, , Dandona, P, 2007
Role of inflammatory mediators in the suppression of insulin receptor phosphorylation
in circulating mononuclear cells of obese subjects.
Diabetologia 50:278–285
Return to RESVERATROL
Return to SHAKLEE STUDIES
Since 2-05-2011
| Home Page | Visit Our Sponsors | Become a Sponsor |
Please read our DISCLAIMER |