|
|
The Diencephalon The Telencephalon The Limbic System The Visual System The Consciousness System The Posterior Fossa Level The Medulla Oblongata The Pons The Midbrain The Cerebellum The Spinal Levels Functional Anatomy Normal Reflex Activity Diagnostic Considerations The Peripheral Levels The Peripheral Nerves The Peripheral Plexuses The Posterior Rami Muscle Histology Clinical Considerations BibliographyChapter 4
The Horizontal Neurologic Levels
From R. C. Schafer, DC, PhD, FICC's best-selling book:
“Basic Principles of Chiropractic Neuroscience”
Second Edition
The following materials are provided as a service to our profession. There is no charge for individuals to copy and file these materials. However, they cannot be sold or used in any group or commercial venture without written permission from ACAPress.
All of Dr. Schafer's books are now available on CDs, with all proceeds being donated
to chiropractic research. Please review the complete list of available books.
Overview Tone The Supratentorial Level
Chapter 4: The Horizontal Neurologic Levels and Related Clinical Concerns
The reader should keep in mind that the various aspects of the nervous system as described in this manual (eg, longitudinal and horizontal systems) are only reference guides that are visualizations of a patient's nervous system in the upright position. They can be likened to the lines of longitude and latitude on a globe of the earth.
From a basic perspective, it has been previously described that a nerve is a medium in which energy is expressed in various frequencies and amplitudes and directed by various external, internal, and coordinated cause-and-effect processes. The sum of frequencies and amplitudes of a nerve can be stated in terms of nerve "tone." There are ranges of normal tone, and if these ranges are consistently subnormal or abnormal, the nerve can be said to be acting in an unhealthy manner for the situation at hand; ie, inhibited (blocked, suppressed, raised-threshold) or irritated (overactive, hypersensitive, lowered-threshold).
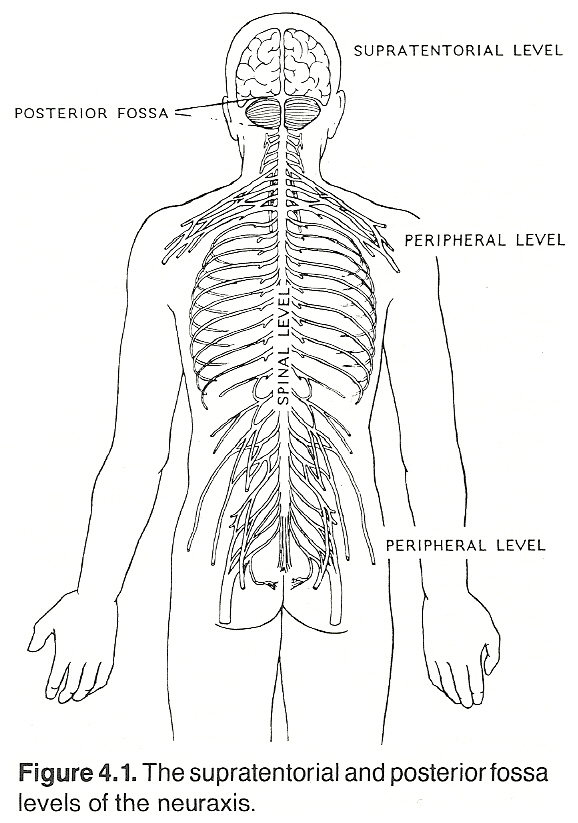
The neural structures located above the tentorium cerebelli are those of the supratentorial level, all which are derivatives of the primitive telencephalon and diencephalon. The major systems at this level are the cerebral hemispheres, the diencephalon, the telencephalon, the limbic system, and central portions of the optic and olfactory nerves.
The Limbic System
Memory
(1) Anterograde amnesia, for events occurring subsequent to the precipitating factor. Some subdivisions are often made; eg,
(a) lacunar amnesia, a patchy loss of memory for isolated events; and
(2) Retrograde amnesia, for events occurring prior to the precipitating factor. Agnosia, astereognosis (tactile memory loss), and certain forms of alexia, agraphia, auditory aphasia (word deafness), and visual aphasia (word blindness), can be considered specific types of amnesia.
Most memory disorders arise within the frontal and temporal lobes. The cause may be gross tissue destruction, a lesion in the association cortex, local cellular changes in the reticular-activating or limbic systems, neurochemical in origin, a RNA synthesis defect, in the neurotransmitters, or in the protein molecules. Infrequently, an obscure low-pressure hydrocephalic condition is involved. It may be subclinical in expression. In overt acute states, fever, headache, nausea, vomiting, and focal signs may be associated.
Olfactory Disorders. The patient's sense of smell can be tested with various unlabeled solutions or oils such as cinnamon, peppermint, clove, menthol, vanilla, wintergreen, etc. See Chapter 8.
(1) remote events such as childhood residence, employment history, names of siblings, etc;
Memory deficits can occur in any of these three areas. They are most prominent in Alzheimer's dementia, amnesia, encephalitis, lesions of the hippocampus-fornix-mamillary body area, temporal lobe lesions, and the Wernicke-Korsakoff syndrome.
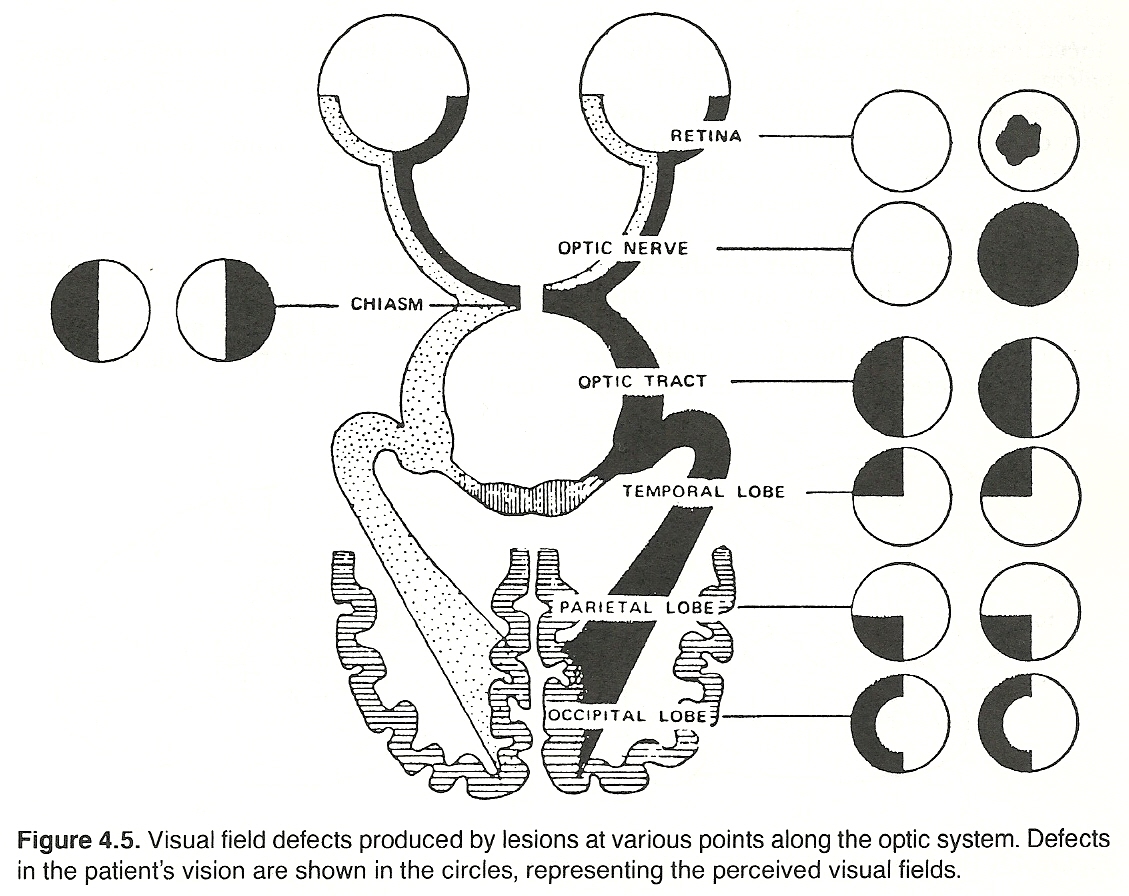
Optic nerves. These tracts of axons extend from each retina, exit through the optic foramen, and unite to form the optic chiasm.
The optic chiasm. The two optic nerves join at the optic chiasm. The fibers decussate within the chiasm in such an arrangement that the nasal portion from each retina crosses to the opposite side and the temporal portion from each retina remains uncrossed. Thus, lesions in the central chiasm express as binasal hemianopsia; lesions in the lateral portions of the chiasm (rare), manifest as bitemporal hemianopsia.
Optic tracts. Because of the decussation that occurs in the optic chiasm, fibers from the nasal retina of the left eye join with fibers from right temporal retina to form the right optic tract. Fibers from the nasal retina of the right eye join with fibers from left temporal retina to form the left optic tract. Each optic tract courses posterolaterally and terminates in the respective lateral geniculate body. A few fibers, however, leave the tract before this and are projected to the pretectal area of the brain to form the afferent limb of the light reflex.
Lateral geniculate bodies. Each lateral geniculate body receives the fibers of the optic tract and projects them, following synapse, via the geniculocalcarine tract. In this manner, the entire left visual field is projected to the right hemisphere, and the right visual field is projected to the left hemisphere.
Unilateral lesions of the optic tract, lateral geniculate bodies, or optic radiations manifest as homonymous defects in the field of vision of one or both eyes, depending on whether the lesion is partial or complete. Lesions of the optic tract anterior to the optic chiasm produce a defect of vision in only one eye.
Optic radiations. The fibers of the diencephalic geniculocalcarine tract extend through the retrolenticular aspect of the internal capsule as optic radiations. The posterosuperior fibers of the radiation course the parietal lobe and terminate in the superior calcarine cortex of the occipital lobe. Unilateral parietal lobe lesions produce a contralateral inferior quadrant defect. The anteroinferior fibers of the radiation extend around and through the temporal lobe and terminate in the inferior calcarine cortex of the occipital lobe. Unilateral temporal lobe lesions produce a contralateral superior quadrant defect.
The occipital cortex. The inverted image of the visual field on the retina is reproduced in a similar topographic fashion in the calcarine cortex of the contralateral occipital lobe; ie, (a) fibers conducting the lower part of the image terminate in the superior calcarine cortex; (b) fibers conducting central (macular) vision terminate in the posterior aspect of the occipital pole; (c) fibers conducting the upper part of the image terminate in the inferior calcarine cortex; and (d) fibers conducting impulses from the peripheral aspects of the retina terminate in the more anterior portion of the calcarine cortex. It is within the cortex of the occipital lobe that the dual images are integrated into one image.
Disorders of the Visual System
The major sensory pathways that provide the CNS with data from its external environment were described in Chapter 3. The highly complex consciousness system is another system. Its ascending portion runs parallel with other primary sensory tracts and provides stimuli for sensory discrimination and levels of wakefulness and attention.
(1) collaterals from all ascending sensory pathways (eg, spinothalamic and lemniscal tracts);
The medial projection area of the reticular formation gives rise to many efferent pathways (eg, thalamus, hypothalamus, cerebellum, and the visceral system). With such diverse input and output, the reticular formation serves as a primary integrator, modifier, and regulator for various levels of the neuraxis.
(1) reticular, along the lateral margin of the thalamus;
Diffuse projections from thalamic nonspecific nuclei to the cerebrum. The ascending thalamocortical reticular activating system offers diffuse projections from the nonspecific nuclei of the thalamus to all areas of the cerebral cortex. These pathways play an important role in modifying levels of consciousness and attentiveness by regulating the electrical activity of the cortex. An individual's degree of consciousness is not determined by a single area in the cortex such as seen in the primary sensory and motor systems; rather, consciousness is regulated by the quality and quantity of exciting, inhibiting, and synchronizing thalamocortical impulses to all areas of the cortex.
(1) local activity,
(1) other cortical neurons, which play a minor role, and
EEG patterns also vary according to which specific thalamic nuclei are stimulated. A rising and falling recruiting response is produced when nonspecific midline nuclei are stimulated. Their potentials are widely projected throughout the cerebral cortex. A diphasic augmenting response occurs when specific sensory nuclei are activated, and their potentials are projected to a specific sensory area of the cortex.
(1) The degree of wakefulness, which is the direct effect of emotions and ARAS input, and the indirect effect of corticoreticular, sensory, and motor stimulation.
Unconsciousness. There are two forms of unconsciousness: pathologic (eg, coma) and physiologic (sleep), and either may be transient or persistent. Transient states of pathologic unconsciousness are commonly associated with syncope, seizures, cranial concussions, and metabolic encephalopathy. The various pathologic states of unconsciousness were described in Chapter 3.
Mental Functions
Presentative vs Representative Powers. While witnessing intellectual processes during daily activities, we can become aware that many relate to perceptions brought to conscious awareness by the specialized peripheral sensory receptors (eg, visible light waves striking the retinae, audible sound waves striking the ear drums, etc). It is in this manner that we become aware of objects within our external environment, here and now. When we come upon an animal, we see the animal, hear the animal, and possibly smell the animal. This ability to perceive sensations of objects present is called the presentative power of the mind and forms the basis of our conception of environmental reality.
Attention
Milstein and associates point out that patients with organic lesions make a large number of mathematical mistakes, but they rarely get excessively disturbed with such errors. These errors are the result of impaired intellectual function and the inability to remember the last number vocalized. Patients with Korsakoff's syndrome, such as seen resulting from the organic brain lesions of chronic alcoholism, habitually give confabulatory responses.
Sleep Patterns. Two distinct cyclic sleep states have been identified: those exhibiting rapid eye movement (REM) and those that do not (nonREM). Each of these states has distinct characteristics. About 50% of adult sleep and 80% of infant sleep are REM sleep. Upon falling to sleep, the first period is nonREM. It lasts about 60—90 minutes and is followed by a REM period, which lasts approximately 5—30 minutes. These cycles, thought to be the result of reciprocal release and inhibition of noradrenergic and serotonin mechanisms in the brain, continued to alternate 4—6 times during normal sleep. Drowsiness, exhibiting a low-amplitude pattern with slight fluctuations in rhythm.
Light moderate sleep, which features (a) frontal region EEG sleep spindles (ie, sinusoidal waves of 10—14 ips); (b) frontal and parietal region V waves (sharp high-amplitude waves); and (c) K complexes (combined spindles and V waves), common to partial arousal.
Deep sleep, characterized by large groups of high- and low-amplitude slow waves.
Arousal from Normal Sleep. The normal physiologic loss of consciousness (sleep state) can easily be reversed by the activation of sensory or motor input to the cerebral cortex. When an individual is gently awakened from the sleep state, the EEG pattern changes from slow smoothly rhythmical waves to fast low-voltage waves. The Circulatory Theory. Because vasculature changes were recorded during sleep, many believed that sleep was induced by a physiologic brain anemia. These changes were attributed to circulatory effects upon the cortex and vasoconstrictive centers of the brain stem.
The Histologic Theory. This theory was based on animal studies that demonstrated CNS nerve cell changes after prolonged stimulation. It was thus hypothesized that sleep was the effect of such changes to allow for cellular revitalization.
The Hypoxic Theory. This theory proposed that sleep was the result of accumulating hypoxia. The supposition was that brain cells during the waking state consumed a greater supply of oxygen than could be fully restored by the circulation. In time, this deficiency was thought to reach a point which induced sleep so that the CNS oxygen supply could be restored during the reduced metabolic state.
The Metabolic Theory. At one time, some authorities held that sleep was induced by the progressive accumulation during wakeful activity of lactic acid, carbon dioxide, and associated metabolic debris.
The Psychobiologic Theory. This theory held that sleep was an instinctive behavior that served to prevent the harmful effects of exhaustion.
The Psychoanalytical Theory. Freud believed that sleep was an escape mechanism that involved a regression to an older, more primitive or infantile state.
The Hypnotic State. The word elements hyp, hypno, and hypna come from the Greek and mean sleep. Thus, the word hypnosis is a confusing misnomer as the state is much better described as a condition of profound highly alert concentration. The eyes need not be closed nor the sleep-resembling state acquired for "hypnotic" suggestibility to occur. This is demonstrated in the contagious atmosphere created by gifted orators and writers such as are used within the communications media, the mass hysteria of mob action, the tics and fads that spread throughout a school, subliminal messages, and the many psychosomatic disorders that are passed from one individual to another.
Dreams
(1) the manifest content, events occurring to the dreamer. The manifest content is usually a transformation, a disguise, and a distortion. It is usually the obvious incidents and circumstances of the dream.
This is usually achieved by having the patient free associate around the dream content and its symbols (repressed desires) until their wish-fulfillment nature and physical expression become apparent. Deeper than either the manifest or latent content there is a more impersonal symbolism that tends to relate the separateness and oneness of life.
Lucid Dreams. While all people dream during sleep, only some dreams can normally be recalled on awakening. La Berge, who heads the Sleep Research Center at Stanford University has been directing investigations into the development of an inexpensive device that can be used by anyone who is interested in inducing lucid dreams (being aware of dreaming while dreaming). La Berge believes that the proper interpretation of lucid dreams can be used to facilitate creative problem solving, enhance self-image concepts, develop higher levels of consciousness, and improve mental, emotional, and perhaps physical health.
Supratentorial Disorders: Basic Considerations
(1) the presence or absence of nervous system function,
As many subjective tests are employed, the practitioner should make every effort to put the patient at ease and to explain unfamiliar procedures beforehand. In some instances, including the tests for general cerebral function, part of the examination can be conducted while taking the case history.
Agnosia
(1) visual, Tactile agnosia is a failure to identify objects by means of tactile and proprioceptive sensibilities when both are normal in that part of the body being tested. Astereognosis is the inability to recognize the form, size, texture, and identity of the object touched, due to sensory impairment at a cortical level. Tactile agnosia is related with lesions of the supramarginal gyrus (parietal lobe) of the left cerebrum.
Visual agnosia is the inability to recognize objects not due to a defect of visual acuity or to intellectual impairment. Visual agnosia is associated with lesions of the lateral visual association areas of the occipital lobe in the dominant hemisphere.
Auditory agnosia is the failure to distinguish different noises or sounds in a patient with normal hearing. It involves lesions in the superior temporal gyrus (area 22) in the dominant hemisphere.
Proprioceptive agnosia is the inability to recognize body parts and relationships due to a lesion within the parietal lobe (posteroinferior regions).
(1) cortical alexia, sensory aphasia due to a left gyrus angularis lesion;
Alexia with agraphia is usually related to an infarction in the middle cerebral artery (parietal lesion). Alexia without agraphia is often the result of an infarction in the posterior cerebral artery or its distribution. Alexia with or without agraphia may be caused by the same disorder that causes aphasia. Stroke is the most common cause. Expressive aphasia. This is a motor aphasia caused by impairment or lack of ability to express thoughts in a meaningful way in speech or writing. It is the inability to utter words although the patient understands spoken and written words and knows what he wants to say. It is impossible for the patient to coordinate the muscles controlling speech because of a lesion or dysfunction of Broca's area. Frequently, both speech and writing (agraphia) are impaired. This aphasic condition is also called aphemia, Broca's aphasia, motor aphasia, or ataxic aphasia. The cause is a lesion of the inferior frontal gyrus, just anterior to Broca's area of the motor cortex.
Receptive aphasia. This is a sensory aphasia caused by impairment in the appreciation of the meanings of both spoken and written words; ie, the inability to comprehend written, spoken, or tactile communications. It is often called sensory or Wernicke's aphasia. The cause is a lesion affecting the posterosuperior temporal gyrus and adjacent angular gyrus of the temporal lobe, disrupting the fibers that receive and integrate central impulses. The larger the area affected, the greater the degree of aphasia.
These two types of aphasia can be further divided into auditory receptive aphasia, visual receptive aphasia, expressive speaking aphasia and expressive writing aphasia. For example, other types of aphasia and related disorders include alalia, amnestic aphasia, auditory aphasia, conduction aphasia, fluent aphasia, gibberish aphasia, nonfluent aphasia, optic aphasia, paragrammatism, and paraphasia.
(1) note spontaneity and fluency of patient's speech;
Other common tests can be used in suspected agnosia, alexia, apraxia, and aphasia. Refer to Table 4.3. Kinetic apraxia is characterized by an inability to perform precise acquired motor movements in the absence of paresis due to lesions involving the precentral cortex.
Ideomotor apraxia is induced by an interruption of pathways between the center for formulation of a motor act and the motor areas necessary for its execution. The patient is unable to perform on command a voluntary movement he or she may know and want to fulfill. Ideomotor apraxia is commonly bilateral and is associated with lesions of the parietal lobe (supramarginal gyrus) of the dominant hemisphere.
Ideational apraxia is characterized by the inability to formulate the conceptual plan for the execution of the components of complex movements. Simple isolated movements can be achieved by the patient with no difficulty. This has been implicated as the result of lesions of the dominant parietal lobe and/or corpus collosum.
Constructional apraxia is a condition that features a loss of visual guidance, impairment of the visual image, and disturbance of revisualization. The patient is unable to reproduce simple geometric figures by drawing or by the arrangement of blocks. As a rule, the patient is unaware of the inability to perceive spacial relationships. The condition is associated with a lesion or interruption of pathways between the occipital and parietal cortex.
(1) unilateral muscular development may be a factor in postural analysis,
The left hemisphere is dominant in a right-handed person, and vice versa. According to Chusid, the dominant (major) hemisphere is mainly concerned with verbal, linguistic, arithmetic, calculating, and analytic functions.
(1) affective disorders,
Cerebral dominance does not become established until about the 3rd or 4th year of life. Some physiologists have related dominance to cerebral asymmetries. Other psychologic studies have concluded that males tend to have a predominantly dominant hemisphere, while females tend to use both hemispheres more equally during conscious activities. Generalized differences in outlook between the sexes are sometimes attributed to this difference.
Gerstmann's Syndrome. This clinical picture strongly suggests parietal dysfunction of the patient's dominant hemisphere. Stroke is usually the cause. Gerstmann's syndrome is a symptom complex consisting of
(1) acalculia, the inability to calculate;
This area contains all structures within the cranium that are situated above the foramen magnum and below the tentorium cerebelli. The major structures are the brain stem (medulla, pons, midbrain), proximal portions of cranial nerves III XII, and cerebellum. The medulla is derived from the metencephalon; the pons, the metencephalon; and the midbrain, the mesencephalon. Those portions of the cranial nerves that are located outside the cranial vault are considered in this text to be part of the peripheral nervous system.
As an overview, the major systems of concern are shown in Table 4.6.
The Medulla Oblongata
Basic Considerations
The Glossopharyngeal Nerve (Cranial IX). The glossopharyngeal is a mixed motor and sensory nerve that supplies the pharynx, posterior taste buds, parotid gland, and nociceptors of the middle ear. Its motor fibers arise from the nucleus ambiguus, project through the lateral medulla, and then pass with the sensory fibers. The sensory fibers arise in the superior and petrous ganglia, pass through the jugular foramen, and enter the medulla just behind the facial nerve.
Cardiac Center. This center has groups of bilateral cells in the medulla at the level of the nucleus of the vagus nerve. Fibers from this center accompany those of the vagi to the heart and unite with branches from the thoracic sympathetics to form the cardiac plexus, which envelops the ascending aorta and its arch. Vagal inhibitory and sympathetic accelerating fibers are true antagonists in terms of heart rate.
Cranial Nerves IX XII. See Table 4.8.
The Glossopharyngeal Nerve. Lesions of the glossopharyngeal nerve commonly arise as glossopharyngeal neuralgia, Bonaire’s syndrome, Vernet's syndrome, or tympanic branch neuralgia of Jacobson's nerve. Symptoms of glossopharyngeal involvement include slight dysphagia and possible ear pain. General signs include an inhibited gag reflex, deviation of the uvula to the well side, loss of pharyngeal constriction during an "ah" test, diminished taste in the posterior third of the tongue, hypersalivation, tachycardia, and possibly associated nystagmus.
The Pons
Basic Considerations
The Trigeminal Nerve (Cranial V). The three-branched trigeminal nerve contains a large sensory and a small motor root. It is the chief sensory nerve of the face and is distributed to the skin of the face, ear, jaw, tongue, gums, and teeth. Its motor fibers supply the muscles of mastication. The sensory root, which contains proprioception fibers from the TMJ and teeth sockets, arises from the gasserian ganglion in the lateral aspect of the cavernous sinus, passes posteriorly between the superior petrosal sinus and the tentorium, penetrates the middle cerebellar peduncle, and then enters the pons.
The trigeminal, abducens, facial, and acoustic nerves may be damaged in the pons or anywhere along their course to the periphery.
The Trigeminal Nerve. Lesions of the trigeminal most frequently manifest as tic douloureux, paratrigeminal syndrome (Reader’s syndrome), or auriculotemporal nerve syndrome. Symptoms of trigeminal involvement include anosmia (from nasal dryness), corneal anesthesia, hearing loss, numbness of upper face, pain, and possibly paresthesia. The signs of a trigeminal lesion include dental cavities, facial ulceration, herpes simplex, jaw deviated laterally, inability to chew, masseter wasting, loss or inhibited reflexes (sneeze, lid, conjunctival, corneal, and jaw jerk), and neurokeratitis.
Basic Considerations. The midbrain is directed upward and forward, and its major structures are: A posterior portion (tectum), which contains four rounded eminences (corpora quadrigemina) arranged in pairs that hold important mediating centers and nuclei concerned with reflex coordination. The superior colliculi extend laterally to the lateral geniculate body, and the more prominent inferior colliculi project to the medial geniculate body. The former is associated with visual reflex activity; the latter, auditory reflex activity.
An intervening passage or tunnel, the cerebral aqueduct (aqueduct of Sylvius) that joins the 3rd and 4th ventricles.
An anterior portion, which contains a pair of cylindrical bodies (cerebral peduncles) that are made up largely of the descending and ascending fiber tracts from the cerebrum, cerebellum, medulla, and spinal cord.
The Oculomotor Nerve (Cranial III). The oculomotor nerve is a motor nerve to all ocular muscles except the lateral rectus and superior oblique. It contains fibers originating from the Edinger-Westphal nucleus. When activated, the pupil constricts and the eyeball rolls down and lateral. The nerve exits the brain on the medial side of the cerebral peduncle. It runs behind the posterior cerebral artery, anterior to the superior cerebellar artery, and lateral to the basilar artery. It then passes anteriorly in the cavernous sinus and exits the skull via the superior orbital fissure.
The Oculomotor Nerve. Inhibiting lesions involving the oculomotor nerve affect the patient's ability to look nasally, up and nasally, up and temporally, and down and temporally. The pupil will be constricted in irritative lesions and dilated in nerve blocks.
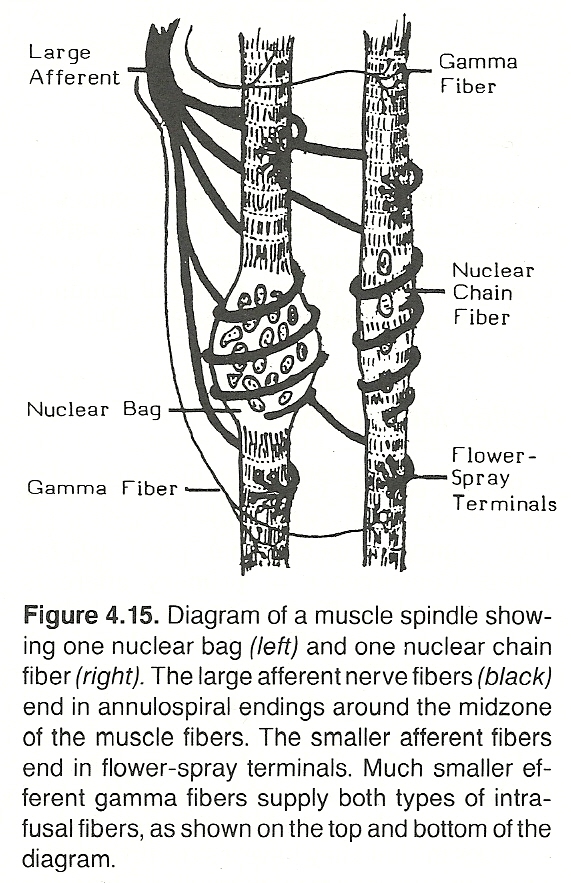
The Cerebellum
Golgi tendon organs are embedded in the musculotendinous area to protect against overstretch. Each Golgi organ is penetrated by one or two terminal sensory fibers, and each communicates in series with several muscle fibers belonging to different motor units.
Reactions to Chiropractic Adjustments. A dynamic chiropractic adjustment, using a high velocity and low amplitude thrust, designed to increase the distance between muscle origin and insertion would quickly elongate the contracted muscle fibers and encourage relaxation via the Golgi tendon organ reflex. On the other hand, slow intermittent manipulation or traction would encourage resistance via the stretch reflex.
(1) cutaneous A-delta and C afferents and
The usual response is a quick ipsilateral flexion activation and extension inhibition (reciprocal innervation) withdrawal of the irritated limb, for example, and contralateral extension activation and flexor inhibition reaction (crossed extension) in the contralateral limb.
(1) those acting upon the neck attempt to keep the head oriented in respect to the body and
These and other righting reflexes are described in Table 4.14.
Spinal Cord Injuries
Concussion of the Spinal Cord. Immediate signs of cord concussion are usually not manifested in mild or moderate injuries; but weeks later, lower extremity weakness and stiffness may be experienced. It takes time for nerve fibers to degenerate. Deep reflexes become exaggerated, and originally mild sensory, bladder, and rectal disturbances progress. The picture is cloudy, often mimicking a number of cord diseases (eg, sclerosis, atrophy, syringomyelia). Life is rarely threatened, but a prognosis of full recovery should be well guarded.
Thus, in a case of scoliotic deviation without body rotation toward the convex side, signs indicating undue tension within the vertebral canal should be sought. It also should be noted that atlanto-occipital, atlantoaxial, and coccygeal disrelationship with partial fixation place a degree of traction upon the cord, dura, and dural sleeves in flexion-extension and lateral bending efforts.
The peripheral levels contain those neurologic structures exterior to the skull, vertebral column, and sacrum; and at each horizontal level, the sensory, motor, visceral, and vascular systems are represented.
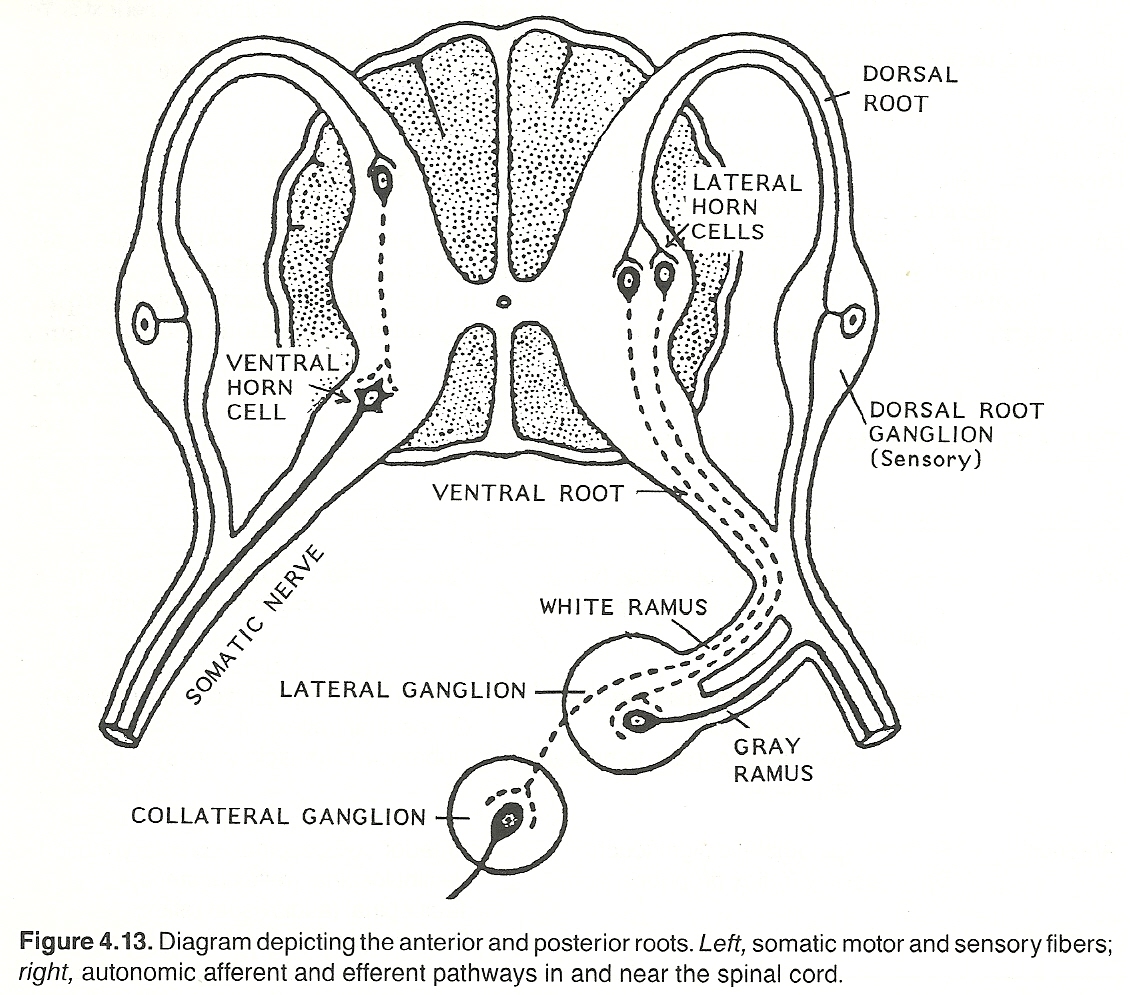
(1) the rami communicantes,
The fibers of the anterior primary division are most prominent in their formation of the cervical, brachial, and lumbosacral plexuses. The usually smaller posterior primary division consists of a medial branch and a lateral branch. The medial branch is sensory for the most part. The lateral branch is primarily a motor nerve, but numerous sensory and autonomic fibers are present.
Significance of Tenderness. Tenderness over the rami of the posterior division has particular significance in the palpatory examination of the spine. The sensory distribution runs in a zone extending from the posterior scalp to the coccygeal area and then laterally to the greater trochanter.
Muscle Histology
(1) the number of nerve fibers employed in carrying stimuli to the muscle fibers,
Besides the above seven factors, the mechanism of the generation of motor nerve impulses in the brain is so susceptible to environmental and psychologic influences that the absolute strength of stimuli (number of nerve fibers and rate of nerve impulses) in two similar situations is unpredictable; eg, motivation influences work output. Different people respond differently to an identical stimulus.
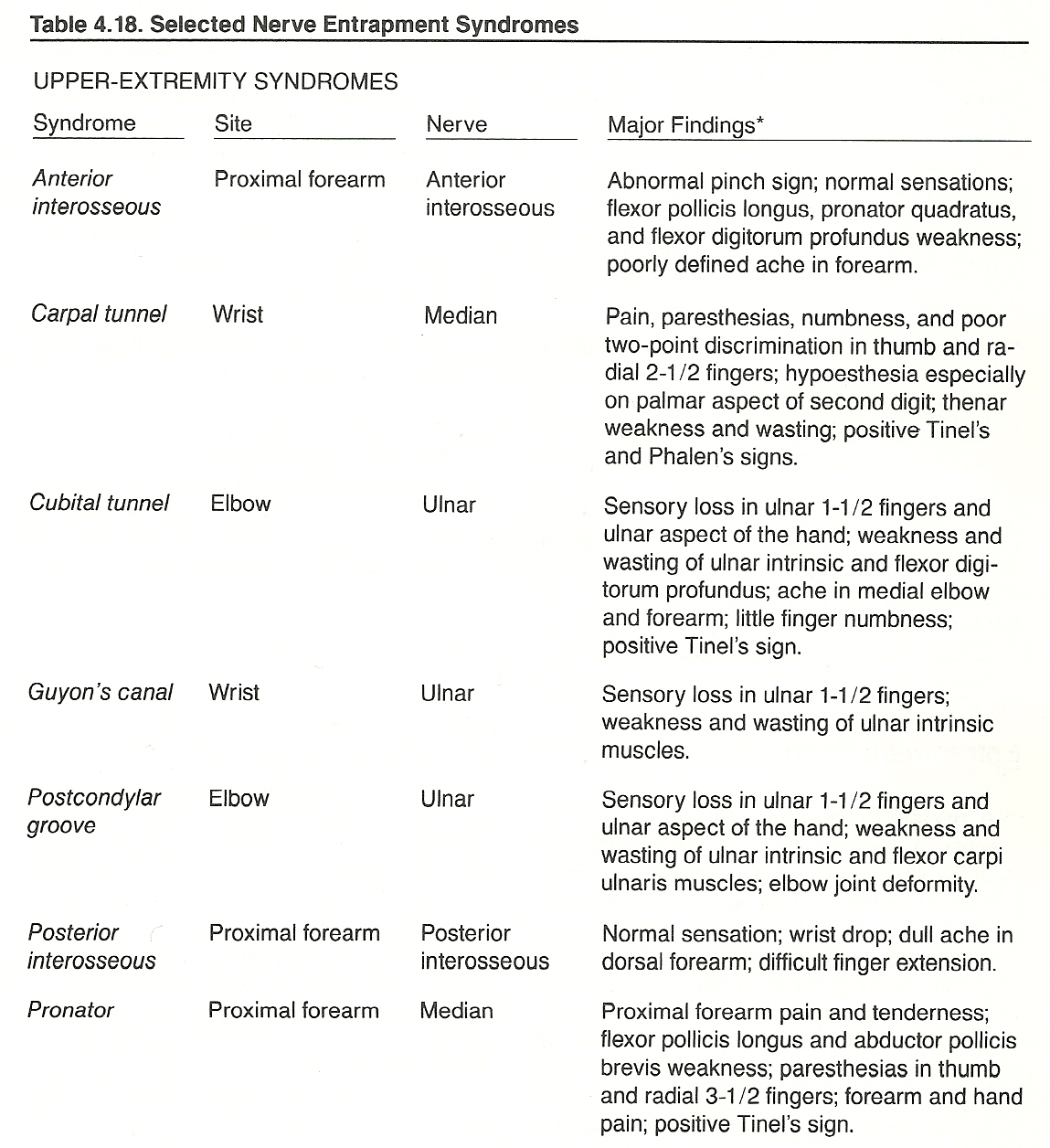
Focal Disorders. Focal lesions are usually caused by trauma, entrapment, or an impinging mass. Nerves that pass through or near joints and dense soft tissues are particularly vulnerable to injury. Vertebral subluxation, carpal tunnel, tarsal tunnel, and other entrapment syndromes are common examples. Some nerves are subject to trauma by being superficially exposed; eg, the peroneal nerve curves outwardly around the head of the fibula at the knee and thus is especially vulnerable to "clipping" injuries in football.
(1) loss of most sensations distally;
Axolemmal Blocks.
This chapter describes the basic functional anatomy and clinical considerations of the horizontal aspects of the supratentorial, posterior fossa, spinal, and peripheral levels of the nervous system.
OVERVIEW
Such systems do not exist physically, but they do serve as excellent mental grid-like tools (viewpoints) during localization and areas in which and from which relationships can be described. For example, although the longitudinal systems take a general vertical course within the spinal column there are numerous alterations and they actually become horizontal when decussating. While the horizontal levels are spatially placed in and extend from the CNS in a general segmental manner, they soon take a widely diffuse course as they project toward their destinations. Thus, references to longitudinal and horizontal levels are just general viewpoints.
It is helpful for study purposes to isolate the body into certain systems, as described above, organize systems into organs, organs into tissues, tissues into cells, and cells into their components. However, we should keep in mind that, physically and functionally, there is only one integrated body and it is more than the sum of its parts. And even the body cannot be thought of as truly separate from its external environment. Although we may do this for study purposes, it is a limited viewpoint.
The human body is the most intricate and efficient electric instrument in the universe. In a state of perfect health, every organ, tissue, and cell functions in perfect unison, one with another. The primary regulator of such activity lies within the CNS. Its higher component, the brain, is much more than a conglomerate of relay stations. It is a highly evolved data gatherer, data processor (selector and organizer), labeler (identifier), transformer, and transmitter of energy.
We do not actually "see" with our eyes, "hear" with our ears, "taste" with our tongue, "smell" with our nose, or "feel" with our fingertips. Our specialized receptor organs are only peripheral sensors for visual, sound, liquid, gas, and pressure stimuli by which impulses are sent centrally so that we can be aware of our external environment.
It is primarily within the brain where all types of nerve impulse frequencies are transformed from input frequencies and amplitudes to selected (consciously or unconsciously) output frequencies and amplitudes. It is within the brain where sensations are transformed into musculoskeletal dynamics and where the stimuli from light, sound, heat, touch, thought, and other action potentials are each in their turn transformed into other frequencies, each influencing body function (specifically and holistically) in either a beneficial or destructive manner.
TONE
When nerve tone reflects itself upon an effector organ (eg, muscle, gland, vessel), a subservient body part is said to be expressing a certain tone. Such descriptors as spasticity and flaccidness in reference to muscles, parasympathicotonia and sympathicotonia in reference to the autonomic nervous system, or anxiety and depression in reference to psychologic states are just a few selected labels to describe particular states of overall voluntary muscle, autonomic, and psychic tone. Thus, in this context, the sum tone of the individual can be considered to represent the general health status of the individual.
In capitalized letters on the title page of what is generally believed to be his first book, Text-Book of the Science, Art and Philosophy of Chiropractic, D. D. Palmer, the father of the science of chiropractic, stated, "FOUNDED ON TONE." On page 7, he continues, "Life is the expression of tone. In that sentence is the basic principle of chiropractic." He went on to state that "Tone is the normal degree of nerve tension. Tone is expressed in functions by normal elasticity, activity, strength and excitability of the various organs, as observed in a state of health. Consequently, the cause of disease is any variation of tone ...."
THE SUPRATENTORIAL LEVEL
The Diencephalon
The structures of the diencephalon, all of which are located between the cerebral hemispheres and the midbrain, surround the 3rd ventricle. The major structures of the diencephalon are the thalamus, hypothalamus, and epithalamus.
The Thalamic Body
The thalamus of each hemisphere is the largest structure in the diencephalon. Its prominent medial portion is called the pulvinar; its lateral oval swelling is called the lateral geniculate body. The centroposterior aspect of the thalamus is the main body of the thalamus. Its nuclei, arranged in anterior and posterior tiers adjacent to the lateral wall of the 3rd ventricle, serve as relay stations for the several ascending and descending tracts that were described in Chapter 3.
The Anterior Thalamus
That portion of the thalamus which lies between the thalamic body and midbrain is the anterior thalamus. It contains nuclei and passing tracts associated with the control circuits of the basal ganglia and the motor system. The subthalamic nucleus is the major nucleus of the anterior thalamus, and a lesion here produces hemiballismus.
The Hypothalamus
The hypothalamus is considered by many to be the central homeostatic core of the autonomic nervous system. It serves as an integrater and regulator of consciousness, defensive emotions (eg, fear, rage), appetitive behavior (eg, hunger, thirst, sex drive), water metabolism, sleep, body temperature, endocrine and cardiovascular functions, and blood osmolality.
The hypothalamus is below and forward to the anterior thalamus and separated from the thalamus by the hypothalamic sulcus. Its major structures are the hypothalamic nuclei in the walls of the 3rd ventricle, tuber cinereum, and mamillary bodies. The hypothalamic nuclei and several tracts are part of the limbic system. Connections are made with all areas of the brain stem and cerebral hemispheres. Although the pituitary gland is not considered part of the hypothalamus, its stalk (infundibulum) attaches to the tuber cinereum. Some authorities consider the optic chiasm a part of the hypothalamus.
The Epithalamus
The epithalamus lies behind the thalamic body, above the anterior thalamus in the posterior wall of the 3rd ventricle, forming part of the roof of the diencephalon. It contains the posterior commissure and pineal gland. The latter secretes melatonin and endorphin containing hormones. The pineal gland begins to calcify after the age of 20 years. It then can serve as a central landmark, during roentgenography, that may be shifted by a space-occupying lesion.
The Telencephalon
The telencephalon is derived from lateral evagination of the rostral part of the embryonic neural tube. The major structures of the telencephalon are the basal ganglia, the subcortical white matter, and the cerebral cortex. These structures fill the entire cranial vault above the tentorium cerebelli and are separated by the falx cerebri.
The Basal Ganglia
The basal ganglia, which surround the ventricles, are three large masses of gray matter (caudate nucleus, putamen, and globus pallidus) located deep within the cerebral hemispheres, anterolateral to the thalamus. The synaptic complexes of the basal ganglia have important function as a control circuit in regulating muscle tone and motor behavior.
It is within the basal ganglia that many lesions of the extrapyramidal tract occur. Parkinsonism and involuntary movements (eg, athetosis, chorea) are typical examples. If the internal capsule becomes involved (eg, hemorrhage), contralateral spastic hemiplegia results.
The Subcortical White Matter
The large mass of white matter beneath the cerebral cortex that surrounds the basal ganglia, contains an array of neuroglia and dense unsynapsing myelinated tracts of afferent (corticipetal) and efferent (corticifugal) projectional neurons that interplay with cortical neurons and such subcortical structures as the basal ganglia, thalamus, hypothalamus, red nucleus, brain-stem reticular formation, cerebellum, and spinal cord (eg, direct activation pathway). This white matter also contains long and short association neurons, which connect ipsilateral hemispheric areas. Short association fibers (eg, subcortical) connect adjacent gyri, while the long fibers connect widely separated areas (eg, the cingulum, fornix, and fasciculi uncinate, arcuate, superior longitudinal, inferior longitudinal, and occipitofrontal).
Numerous transverse commissural neurons are also found in the subcortical white matter. They connect homologous areas of the two hemispheres. The largest commissural mass is the broad corpus callosum, which forms the roof of the lateral and 3rd ventricles.
The Cerebral Cortex
The outer neural surface of the five lobes of the brain consists of a relatively thin mantle of gray matter surrounding the subcortical white matter. It has millions of pyramidal and stellate (granule) cell bodies, organized into approximately six laminated layers, and associated vertically and horizontally running axons, dendrites, and blood vessels.
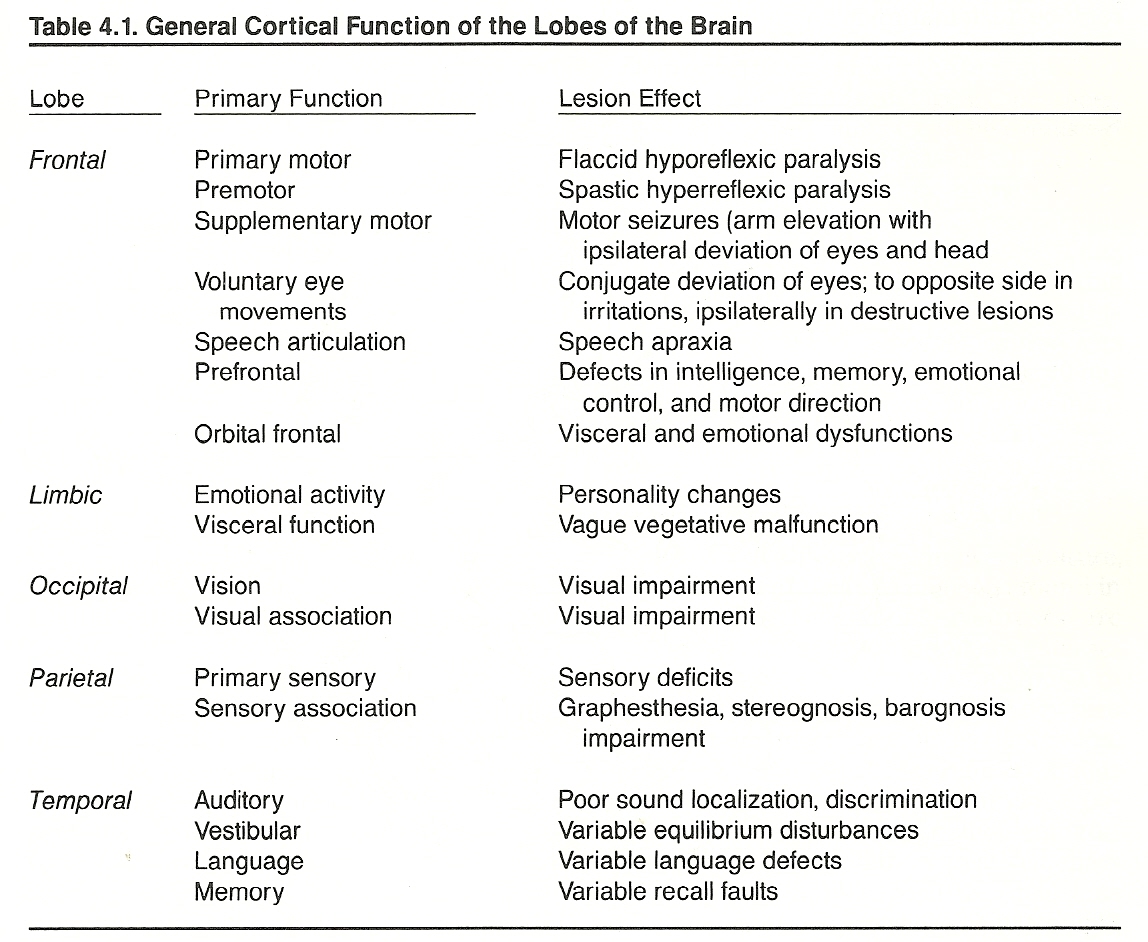
Specific functions are shown in Table 4.1. As Guyton points out, we know less about the mechanisms of the cerebral cortex than we do almost all other parts of the brain, though it is by far the largest portion of the nervous system.
The functionally diverse limbic system includes the limbic lobe of the cerebral cortex, thalamic and hypothalamic areas, and olfactory areas. These structures, arranged in a ring and located deep on the medial surface of the brain, serve to integrate and/or mediate olfaction, behavioral, emotional, memory, and visceral activities. Areas within the frontal lobe (orbital and anterior), medial temporal lobe, and medial parietal lobe are part of the limbic system.
The limbic system as a whole is sometimes referred to as the visceral brain. Specific areas within the limbic system have been isolated that play an important role in eating, sleeping, wakefulness states, gastrointestinal and anterior pituitary function, body water regulation, and sexual activities. The limbic system has also been shown to play a role in behavioral modification (reward-punishment reactions) and some modification of pain. However, limbic lesions usually manifest clinically as olfactory, memory, and/or behavioral alterations.
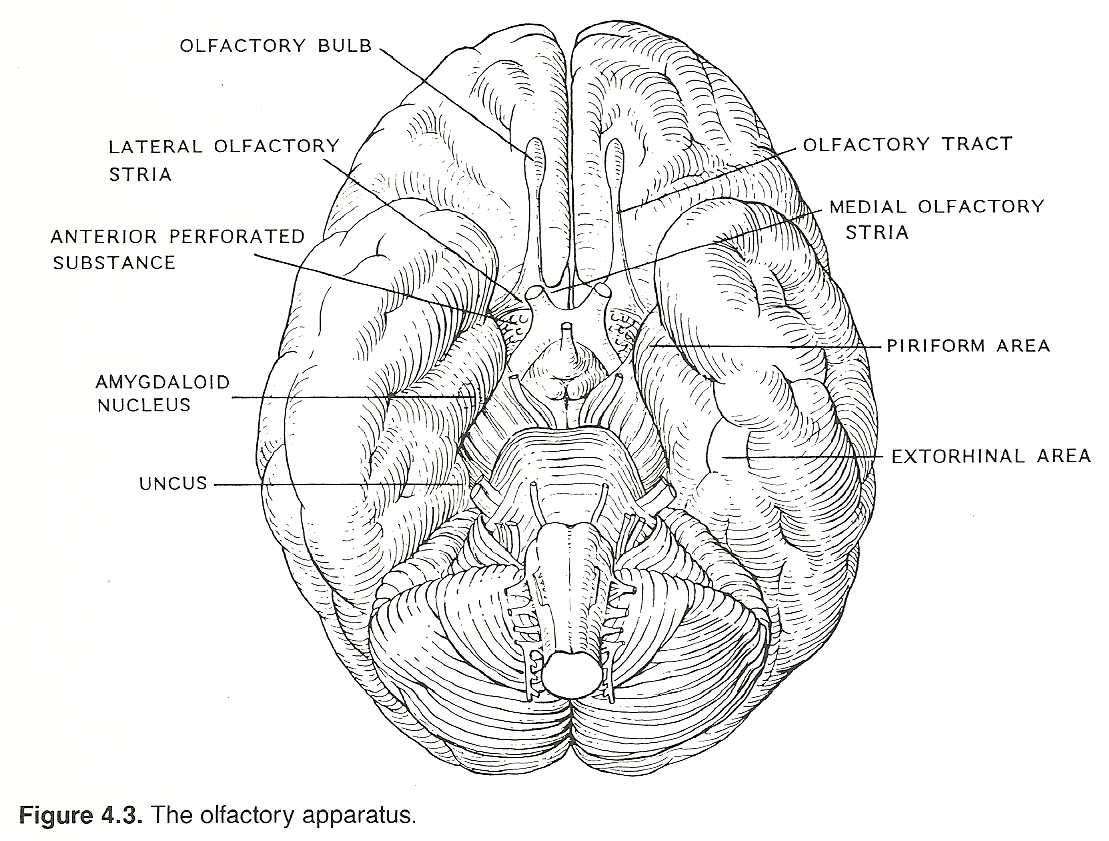
The Olfactory Apparatus
The olfactory nerve is one of two systems solely contained at the cranial peripheral and supratentorial levels. The other is the optic nerve. Both these systems are structured so that they transfer only one type of stimuli. The nose cleans, warms, and humidifies over 500 cubic feet of air every day.
The sensory receptors of cranial nerve I are embedded in the superior nasal mucosa. They are sensitive to gaseous stimuli that are perceived as the sensations of smell. Impulses are conveyed by many small fibers through the cribriform plate to cell bodies in the olfactory bulb, which lies at the anterior aspect of the olfactory sulcus on the orbital surface of the frontal lobe. On reaching the bulb, the fibers divide.
A medial tract extends to the medial basal portion of the frontal lobe. A lateral tract is projected to the medial portion (piriform) of the temporal lobe, concluding in the uncus (chief olfactory area) and amygdala. From these structures, a network of small links is made with other structures of the limbic system. One example of the complex reflex connections is that it is impossible to sneeze and keep one's eyes open at the same time.
Memory is a complicated process of labeling, organizing, classifying, storing, retrieving, and transferring acquired and genetic sensory impressions. These sensory impressions may originate extrinsically from the environment or intrinsically by the formulation of images, concepts, and patterns. Although memory is the prerequisite for learning, it is separate from the "thinking" process. This is decidedly demonstrated in various forms of amnesia and agnosia exhibited by otherwise intelligent individuals.
Memory can be divided into several groups such as visual memory (descriptions of things out of sight), gustatory memory (descriptions of foods not present), olfactory memory (descriptions of odors not present), verbal memory (description of thoughts spoken previously), and motor memory (the repetition of a physical act done previously). Typical examples of inherited memories are instinctive behaviors and the basic urges.
Although some structures are more important for memory than others, there is no isolated area of the brain that regulates the recall of past impressions. Memory stations appear to be dispersed throughout each hemisphere, and data are readily transferred from one hemisphere to the other. Some studies have indicated that areas of the prefrontal cortex store short-term memories and the hippocampus of the temporal lobe stores both complex short-term and long-range memories.
Amnesia
The word amnesia refers to the pathologic or physiologic loss of memory. It may be transient or permanent. The two major types are:
(b) retroanterograde amnesia, a situation where past events are referred to the present and vice versa.
Orientation
Orientation serves as the foundation for most memory and intellectual processes. Inaccurate knowledge of one's specific orientation with time, place, person, or situation is often reflected in a patient's answers during an interview. It may be exhibited in altered history facts on return visits, the inability to follow simple instructions, the inability to sketch the route taken to the office, the inability to judge the approximate time necessary to conduct a simple task, the inability to judge distances to objects in the environment, etc.
Disorders of the Limbic System
Memory Alterations. If the patient's level of consciousness and intelligence has been judged adequate, memory can be evaluated for
(2) recent events such as current address, telephone number, spouse's age, etc; and
(3) immediate recall such as counting seven numbers forward from 57 and counting 5 numbers backward.
Behavioral Disorders. In every culture, evidence of mental illness has been found in the earliest of civilizations. Neuroses are emotional maladaptation problems, characterized by anxiety resulting from some unresolved unconscious conflict(s). They also feature phobic reactions and/or depression. The general term psychoses refers to major mental disorders that grossly impair a patient's capability to meet daily demands. They are characterized by regressive behavior, diminished impulse control, delusions and hallucinations, inappropriate moods, schizophrenia, and often violent antisocial behavior. These often complex disorders are the subject of psychologic and psychiatric studies, thus they will only be referred to in this text in passing.
The Visual System
The eyes are remarkable organs. They can perceive more than 1 million simultaneous visual impressions and can discriminate among nearly 8 million graduations of color. Vision accounts for 90% or more of all sensory impressions.
The neural circuits for all direct optic impulses are restricted to the peripheral and supratentorial levels (diencephalon and telencephalon). The ocular muscles are derived from mesoderm; the other nonneural portions of the eye (cornea, sclera, iris, lens, and humors) are derivatives of embryonic ectoderm.
The Peripheral Neural System
The sensory organs of the retina (photoreceptor rods and cones) are the peripheral receptors of the visual system. These receptors are activated by a narrow band of electromagnetic spectrum that is between infrared and ultraviolet wavelengths.
The 100-million low-threshold, poorly discriminative, monochromatic rods in each eye are responsible for low-intensity (scotopic) night vision and are found in greatest numbers in the peripheral retina. Rods are absent in the fovea. The higher threshold, more discriminative, trichromatic cones serve in high-intensity (phototopic) daylight vision, visual discrimination, and color vision. They are found in greatest numbers at the macula.
Central images are projected upon the posterior poles of each eye. Images of the left field of vision are projected on the left nasal and right temporal halves of the two retinas, and vice versa. In addition, the visual image striking the retina is inverted so that the upper portion of the visual field strikes the lower portion of the retina and the lower portion of the visual field is impressed upon the upper aspect of the retina.
A constant image projected upon a specific area of the retina would rapidly lead to receptor adaptation due to the sustained stimulation. Thus, the eyes normally make rapid minute continuous movements (microsaccades) that mistakenly appear as a fixed gaze.
When light rays strike the rods and cones, a breakdown of visual pigments occurs that leads to the set up of receptor potentials. Dendrite impulses are conveyed to peripheral first-order axons (bipolar cells) that synapse with more distal intermediate neurons and then to further distal third-order neurons (ganglion cells). The latter neurons provide the surface of the retina and come into contact with the vitreous humor.
The ganglion cells send unmyelinated axons from the peripheral retina to the cup-like optic disc, the axons become myelinated at the disc and then exit the eye as the optic nerve. Ophthalmoscopy will reveal lesions of the distal optic nerve such as atrophy, neuritis, and papilledema.
The Central Neural System
Proximal to the eye, the major visual structures are located in the diencephalon and telencephalon. The diencephalic structures of the eye include:
The telencephalic structures of the eye include:
Fibers originating from the macula are widely distributed in the visual cortex. For this reason, unilateral lesions in the visual cortex will usually spare central vision. Visual hallucinations are commonly reported by patients with visual cortex dysfunction.
Communicating axons from the calcarine cortex to the brain stem provide data for unconscious eye motions. Voluntary eye movements and fixation are directed by the eye centers of the frontal lobes of the cerebrum (area 8). Lesions in this area produce conjugate deviations.
Impairment of the visual system may be the result of retinal disease or a lesion of the optic nerve, optic chiasm, optic tract, or occipital lobe. Loss of vision often accompanies a fracture through the sphenoid involving the orbital foramen.
Complete blindness of one eye invariably indicates a lesion of an optic nerve. Optic neuritis and tumors compressing an optic nerve are the most common lesions causing loss of the direct light reflex. Lesions of an optic nerve are especially noted with optic atrophy, optic neuritis, papilledema, and visual impairment. Thus, the optic nerves should be tested for visual acuity and fields of vision, and directly inspected by observing the condition of the fundi.
Homewood points out that, as the nasal mucosa, preganglionic fibers from T1—T4 which connect with postganglionic fibers of the superior cervical ganglion project to the dilator pupillae, part of the levator palpebrarum, the orbital MIller muscle, and the arteries of the eyeball and adnexia. Thus, upper cervical and upper thoracic subluxations may cause or encourage a wide number of motor and vasomotor syndromes of the eye.
These fibers originating from T1—T4 and the superior cervical ganglion supply many of the same tissues innervated by the oculomotor, trochlear, abducens, auditory, and spinal accessory cranial nerves. For this reason, visual, auditory, and equilibrium disturbances are frequency associated with whiplash injuries of the cervical and upper thoracic spine. Less frequently, but not rarely, vagal and phrenic syndromes may be related.
The Consciousness System
In 1909, Brodmann mapped the cerebral cortex on the basis of cortical cytoarchitectual patterns. He numbered 47 distinctive areas of varying thickness. Since then, over 200 specific areas have been isolated, but Brodmann's original areas remain the basic reference (Table 4.2).

Both systems, the primary sensory and the consciousness, end in the cerebral cortex, but they take different routes. The primary sensory system courses through specific thalamic nuclei and then extends to the sensory cortex. In contrast, impulses of the consciousness system travel over indirect, polysynaptic, multineuronal routes to nonspecific thalamic nuclei and then extend to all areas of the cerebral cortex.
It must be kept in mind when studying the sensory system that the only thing sensory nerves do is transmit impulse patterns. The consciousness system, however, does not interpret such signals as just impulse signals. The interpretation of receptor stimulation is experienced subjectively as feelings of touch, pain, temperature, or movement or as sensations of flavor, smell, sight, or sound. And beyond this, in consciousness, there is some mysterious "something" that witnesses all this and is apart from it. The processes of neural circuitry, convergence, summation, facilitation, inhibition, and divergence explain the "where" and "why" of some phenomena but fall far short of explaining the "what" and "how" of it all.
Anatomical Considerations
The major divisions of the consciousness system are the ascending reticular activating system (ARAS) and the final widespread terminations in the cerebral cortex. The ARAS is an aggregate of neurons that receives and integrates impulses from the sensory system, cerebral cortex, cerebellum, hypothalamus, and brain-stem nuclei and then projects them via modifying nonspecific nuclei of the thalamus to all areas of the cerebral cortex for connection with the sensory, motor, and autonomic systems.
The Major Structures of the ARAS
Portions of the brain-stem reticular formation. The reticular formation (network) has complex clusters of diffuse and poorly organized large and small multipolar neurons and associated interneurons located in the central tegmental portion of the brain stem that extend from the decussation of the pyramids in the medulla to the thalamus.
The lateral receptive area of the reticular formation receives afferent impulses from
(2) the cerebral cortex (corticoreticular, corticospinal, corticobulbar fibers);
(3) visceral afferents originating in the spinal cord and cranial nerves; and
(4) the basal ganglia, hypothalamus, cranial nerve nuclei, and cerebellum.
The ascending projection system. Ascending fibers of the reticular formation are not located in specific tracts. They project diffusely from the brain stem (through the pons, midbrain, and diencephalon) to the hypothalamus and nonspecific nuclei of the thalamus.
Nonspecific nuclei of the thalamus. The nonspecific nuclei of the thalamus are the
(2) midline, near the massa intermedia; and
(3) intralaminar, along the internal medullary lamina.
Physiologic Considerations
Because of their aggregate arrangement and multisynaptic connections, the neurons of the CNS do not function singularly. The electric activity of the cortical aggregates is the summed effect of the cortical action potentials occurring at the time. It is derived from
(2) input from specific thalamic nuclei, and
(3) input from nonspecific thalamic nuclei. This summed effect produces an oscillating wave form of electric activity, rather than the simple spikes witnessed in the peripheral nervous system, that can be measured on an EEG.
EEG Tracings. The EEG tracing reflects the summed synaptic potentials of the dendrites lying near the periphery of the cerebral cortex, and the fluctuations represent the varied excitatory and inhibitory synaptic potentials impinging on the dendrites.
The activity of the cerebral cortex is modulated by two forces:
(2) neurons from the thalamus, which play a major role. Synchronizing "pacemaker" impulses from the thalamus determine the intrinsic resting frequencies of brain waves. Low-frequency thalamic impulses produce a synchronized rhythmic high-amplitude EEG wave pattern, while high-frequency impulses produce an opposite effect; ie, a desynchronized arrhythmic low-amplitude EEG wave pattern.
Consciousness. The faculties of consciousness such as alertness, perception, apperception, attention, memory recall, and the regulation of sleep are the primary responsibilities of the cerebral cortex and its input. These states can be viewed from two aspects:
(2) Content, which is the product of cognitive mental activity occurring in the cerebral cortex.
The three primary attributes of the mind (a functional process) are thinking (intellectual thought development), feeling (the perception of sensation), and choosing (volition, the selection of voluntary action, will). Every mental action has one or more of these attributes as its foundation.
The mind, however, not only receives impressions acting on the peripheral sensors from external objects as here and now, it also can conceive of them during their absence and represent them to itself by mental images and thought forms. It can also relate one or more objects to other objects and to certain circumstantial actions and feelings. If we had at one time come upon a wild animal and the experience made a strong impression, we can remember how the animal looked, what the associated circumstances were, and how we felt at the time. This ability to perceive sensations of objects absent is called the representative power of the mind and forms the basis of simple recall, reflection, and situational memories.
Creativity Powers. Unlike either common presentation and representation of objects and situations experienced, the mind it also capable of imaging objects and situations completely different from any experienced (eg, a horseless carriage, a machine that can fly, a six-headed cow) to meet its fancy and ideals. This creative or inventive form of representative power, which is separate from that derived from sensory impressions, forms the basis of imagination as well as hallucination. With this ability, we can conceive of time, space, cause, effect, beauty, etc. Such conceptions may be suggested by sensed objects, but they are not conveyed to the mind through the senses. This ability is called the originative (intuitive) power of the mind.
Classification and Reasoning Powers. The mental functions described above encompass the abilities to present, represent, and create. Another important attribute is the ability to generalize and classify things, thoughts, and acts. That is, we may perceive certain common qualities, properties, and relations of objects rather that the objects themselves. Through such comparisons, we can group, stereotype, and catalog things such as by their visual impression (eg, color, size), audible impression (sounds), tactile impression (eg, texture, temperature), olfactory impression (smell), taste (eg, salty, bitter, sweet, spicy), proprioception (eg, weight), composition (eg, animal, vegetable, mineral components), function, etc. Such generalizations form the basis of our mental labels, codes, filing systems, and tabulations. They also form the basis of our likes, dislikes, and prejudices.
Rather than classifying certain objects by their like elements, we may reverse the process and analyze (inductively or deductively) the component properties and relations within specific things, thoughts, and acts. While such characteristics are the objects of thought and related primarily to sensible objects, the conceptions are not those presented by the senses. This involves the ability to perceive abstract properties and relationships, and the process is the basis of reasoning. Reason can be defined as the logical process of selecting associated ideas into an organized form. Thus, impressions of sense and discursive notions of understanding must be differentiated to maintain clear thinking.
In addition to its state of alertness, the consciousness system is also influenced by its attention toward or away from certain sensory input. That is, by enhancing or suppressing specific impulse transmission, the consciousness system can focus selective attention on the minute details of certain sources and patterns of input and ignore other stimuli. This is commonly seen when an individual is so absorbed in some type of work that various potentially distracting (but not harmful) environmental stimuli are completely blocked from consciousness. This is commonly witnessed when a person is concentrating on reading a book or watching a highly interesting event.
Attention span can be severely shortened by either physical or psychic mechanisms. It is commonly observed that concentrated effort is greatly limited in disorders associated with pain or fever. Likewise, emotional preoccupations can make it impossible to fix attention on a singular subject, regardless of its long-term importance to the individual. This should not be confused with simple habitual procrastination.
The attention span of a patient with an average IQ or better can be simply tested clinically by the Serial Sevens Subtraction Test (Table 4.3). Patients with severe emotional distress (eg, acute anxiety) or paranoid tendencies frequently become highly threatened by such a test.

Attention and the Alpha State
While it is generally held that the alpha-brainwave state cannot be obtained with the eyes open (because of the superimpingement of visual stimuli), distinct alpha states have been recorded while subjects are in a state of profound abstraction, meditation, imagery, or reverie (daydreaming) with the eyes open. Such a state may also occur when we become a witness to our personal thoughts and actions.
Thus, mental processes can be controlled to such an extent that attention can be focused one-directionally to the point where impulses from the periphery will be ignored. This is not an unusual phenomenon. It occurs whenever we become completely engrossed in a book, a movie, or some other event to the degree that we become unconscious of potentially distracting environmental influences. During wartime, it is not unusual for soldiers to report that they were unaware of their wounds until the battle was over.
Such a mental focus or ability to think of one thing at a time is achieved from the faculty of concentration (intensified attention). It is the basis of all learning and achievement and is regulated by personal need, desire, sincerity, and motivation. We see this process manifesting constructively by watching a child learning to walk, by watching an adolescent learning to drive a car, or by watching someone master a complicated dance routine. We witness this steady focus by watching a child crippled with poliomyelitis perseveringly mold into an adult Olympic athlete or by watching an individual with meager educational, financial, or social advantages slowly attain to high status not because of their advantages, but in spite of their handicaps. We also see this process manifesting destructively by watching the circumstantial fruits of the habitual pessimist. Thus, the faculty of concentration is a dual-edged sword.
Abstraction
A patient's ability to deal with abstract concepts is often impaired in organic mental illness. Psychotic patients and schizophrenics tend to become strikingly literal in their interpretation of speech and fail to understand the meaning of commonly used metaphors, proverbs, and other nonliteral expressions of speech. Thus, testing a patient's ability to understand abstractions is often helpful in differentiating the functional neurotic from the organic psychotic.
Thought Content and Association
Thought content may indicate such abnormalities as severe phobias, obsessions, morbid preoccupations, paranoia, or delusions. The patient with forebrain disease will be unemotional, mildly disoriented, and inattentive to personal details.
Thought association reflects the quality of interconnection between a sequence of ideas. When thought association is not orderly, such as speaking in disconnected sentences or offering an answer that is unparallel to a question asked, schizophrenia should be suspected. In schizophrenia and a few other mental syndromes, this inability to associate appears to be the result of impairment of various interrelated neuronal loops in the brain that serve to interconnect thought processes for an orderly sequence of thinking.
Body reactions indicate how completely the individual has entered a situation. Thus, an emotional response is a total measurement of mind, will, and body (including the endocrine system and all its ramifications). An individual's overenthusiastic emotions may alter one's psychic approach to life. Perception is changed, and values are amended. When under the influence of "hot" emotions, life appears different from that which is led by "cool" intelligence. The fading of an emotion is regretted if the associated feeling is pleasurable.
Gross impairment of judgment is characteristic of an organic lesion or severe psychosis. Minor impairments often signal the early stage of an organic lesion. This is often reflected in poor judgment of personal affairs (eg, personal relationships) rather than in nonpersonal affairs (eg, conceptual differentiation). A cursory estimation of a patient's quality of judgment can be obtained by discussing the patient's plans for the future and contingency plans if the ideal outcome is thwarted by some unanticipated obstacle.
Sleep States
When somatic or mental fatigue arises, the level of consciousness tends to decrease into a transient sleep state. This is not a passive phenomenon; ie, caused solely by decreased input to the cerebral cortex. Rather, it is a dynamic phenomenon and currently thought to be the result of activation of certain hypnogenic areas of the brain. These areas are believed to produce neurochemicals that actively promote inhibited cerebral activity and suppress the ARAS. Two hypnogenic areas have been isolated. One is on the anterior aspect of the hypothalamus, and the other is in the upper medullary and lower pontine portion of the reticular formation. Lesions in these two areas will produce a constant state of wakefulness.
A more or less permanent deep sleep state (eg, coma) results if some of the consciousness system becomes impaired. The most frequent site for such a lesion occurs at the midbrain level of the ARAS, especially at the rostral portion of the brain stem and thalamus, above the trigeminal nerve entry zone. Bilateral lesions affecting the ARAS in the diencephalon produce a state of sustained somnolence with diffuse slow-wave EEG activity. A unilateral cortical lesion will not produce unconsciousness if the other hemisphere is functioning, but unconsciousness results if both hemispheres are impaired.
A number of abnormal sleep states are defined in Table 4.4.

REM Sleep. REM sleep, the sleep state in which dreams appear, features rapid conjugate motions of the eye, an increase in body movements and muscle twitches, fluctuations in respiratory and pulse rates, variations in blood pressure and body temperature, decreased muscle tone, and penile erection. EEG patterns reveal a fast wave similar to that of wakefulness with the eyes open but of much lower amplitude.
NonREM Sleep. NonREM sleep is characterized by a lack of rapid eye motion, fewer body movements and muscle twitches, more stable respiratory and pulse rates, and less variation in blood pressure or body temperature. NonREM sleep can be divided into three distinct EEG patterns:
Insomnia (Sleeplessness). The symptom of insomnia, the inability to sleep, should not be taken lightly. It may be the result of an organic lesion, and this commonly occurs at the midpontine pretrigeminal level, rostal to the trigeminal nerve entry zone and above the medullary and pontine sleep centers. If the lesion is severe, the state of wakefulness will be persistent. Milder forms of insomnia also may be due to other symptoms producing excessive sensory stimuli, a certain therapy, the patient's life-style, or various psychogenic factors.
Cirrhosis of the liver, for example, often produces insomnia because of episodic nocturnal delirium. Overeating late at night may produce a reflux esophagitis, and overdrinking in the late evening may produce an alcoholic gastritis. The nocturia of renal or bladder disease, the palpitations and arrhythmia of heart disease, and the dyspnea of pulmonary disease commonly cause insomnia. Coronary insufficiency frequently manifests only an early sign of nocturnal "heartburn." Any upper airway obstruction (eg, rhinitis, epiglottitis) will produce a fear of not being able to breathe during sleep that may maintain wakefulness.
Hyperthyroidism, anemia of any type, heart disorders, and neurocirculatory asthenia are commonly associated with insomnia because of tachycardia. Brain tumors and inflammations frequently cause wakefulness and somnolence, and a large number of other neurologic disorders are associated with insomnia. The latter would include peripheral hypersensitiveness such as pressure on sensitive trigger points.
Sleep Theories. There have been several theories on the causation of sleep that have now been discarded. They are briefly described below for historical interest only.
Dreams are commonly defined as a more or less coherent train of partial conscious (preconscious) imagery occurring during sleep. They also may be induced in drugged or hypnotic states. It appears that during such sleep states thought residues from daily activities join with unconscious tendencies and memories that have been excluded from consciousness by the repression or suppression of painful ideas and form symbolic images of wishes, fears, or weaknesses. This allows some idea within the subconscious to express itself that would normally not be allowed to do so. These are the common wish-fulfilling dreams of some unsatisfied desire or dreams that reflect from a subconscious conflict.
There are also dreams that can be attributed to physical causes such as environmental noises, exposure to temperature extremes while sleeping, or indigestion. In addition, there are those extraordinary dreams that can be placed under the general category of enlightenment (eg, problem-solving, creative, spiritual).
The content of a dream (ideas expressed in a dream) is held to be of two fundamental types:
(2) The latent content (hidden residual thoughts), which must be revealed by the technique of psychoanalytic dream interpretation.
The main objective of the neurologic evaluation is to determine
(2) the location, type, and extent of lesions (if any), and
(3) the degree to which the healthy portion of the patient's nervous system can be used for rehabilitation. Thus, the standard physical examination should be conducted before a more detailed neurologic assessment so that findings may be integrated to reach a conclusive diagnosis.
The effectiveness of therapeutic progress of a patient is often in direct proportion to the patient's psychologic acceptance of the doctor and the therapy. Kirby points out that it is therefore valuable in patient evaluation to judge the ability and willingness of the patient to follow the physician's orders and to create an atmosphere where the patient will have optimum confidence in the doctor's ability and offer optimal cooperation. Conversely, it is well to recognize certain types of patients who tend to be uncooperative and do not have the frame of mind necessary to achieve optimal results, even when the best diagnostic and therapeutic methods are applied.
When patients are obviously not responding adequately during the history interview according to their background, occupation, and educational experience, a mental disorder should be suspected. During the early stages of mental illness, however, such faults may be extremely subtle.
A patient with a severe consciousness disturbance may require immediate surgical or psychiatric referral until the cause can be determined. Such situations are often urgent, and if the patient is conscious, uncooperativeness is the rule. Specialized experience, brain scans, EEGs, and CSF examinations are frequently required during the diagnostic process. For these reasons, the following descriptions for a conscious patient are brief.
Gross evaluation of the higher centers begins with a general mental and emotional evaluation during the case history process that extends throughout the examination noting gait, carriage, handshake, gestures, facial expressions, grooming, cleanliness, and dress.
Portrayed self-image, mood, judgment, response to simple requests, temporal and spatial orientation, memory, attention span, and language comprehension and articulation also can be generally appraised at this time. Language processing involves listening, comprehending, speaking, reading, and writing skills.
Once all examination data are collected, the practitioner should be able to determine whether the cerebral cortex is functioning adequately at its highest levels of integration.
Typical syndromes likely to involve the cerebral cortex are listed in Table 4.5.

Agnosia is the inability to name, properly use, or comprehend the importance of sensory impressions although the sensory sphere and language capabilities are intact. There are seven types:
(2) auditory (acoustic),
(3) olfactory,
(4) tactile,
(5) gustatory,
(6) digital (inability to count fingers), and
(7) temporal (time duration). Four major types of agnosia are described by Tran/Maori as follows:
Alexia and Agraphia
Alexia is the inability to read (word blindness) because of a central nervous system lesion. Agraphia is the inability to express thoughts in writing because of a lesion in the cerebral cortex.
There are four main types of alexia:
(2) motor alexia, inability to read a word aloud even though the patient understands the word when he sees it;
(3) musical alexia, sensory or optic (never motor) inability to read music; and
(4) subcortical alexia, alexia due to interruption of the connection between the gyrus angularis and optic center.
Aphasia
A normal educated person can understand and communicate in spoken and written language, as well as by gestures and other means. Any disorder that is characterized by receptive and expressive disturbances in the faculty of using symbols and signs to communicate is called aphasia.
Aphasia can be caused by organic neural lesions that affect the cerebral memory mechanisms for language without impairment of cortical or subcortical structures essential for the relay of impulses or the innervation of the speech organs. Although it is difficult to pinpoint the location(s) of such lesions, most researchers suggest that the dysfunction is associated with cerebral damages in the posterior temporoparietal region and Broca's area (pars opercularis and triangularis of the inferior frontal gyrus).
Aphasia is an impairment characterized by defective or loss of expression by speech, writing, or signs, or of comprehending spoken or written words because of a cerebral lesion or dysfunction (usually cortical, rarely subcortical), especially that of the left hemisphere. There are two main types:
Isolated receptive or expressive aphasias are rare. Brain lesions that are large enough to cause speech impairment seldom produce pure effects.
The inability to articulate words properly (dysarthria) is a motor disorder and not an aphasia. Aphasic disorders are usually caused by the same diseases that cause hemiparesis; ie, infarct, stroke, trauma, tumor, aneurysm, infection, inflammatory disease, and degenerative disease. Alzheimer's disease and Pick's disease cause aphasia but not weakness.
Aphasia, agnosia, alexia, and apraxia are closely related. The etiologies of aphasia and agnosia, especially, are similar. The purpose of clinical and laboratory tests should be to confirm or exclude psychic, functional, and organic suspicions of causal relationships.
When aphasia is suspected, Heilman recommends five tests:
(2) test comprehension of spoken and written words;
(3) test ability to repeat sentences;
(4) test ability to name familiar objects; and
(5) test ability to write familiar words. The proper interpretation of these tests will help locate the site of the lesion.
Apraxia
Lesions in the cerebral cortex may involve the inability to perform certain learned complex movements in the absence of paralysis, sensory loss, or disturbances of coordination. Apraxia is the inability to perform purposefully learned motor acts on command though physical ability and willingness are present; ie, motor and sensory impairments are not involved. The fault is within the neural pathways that retain memory of learned movement patterns so that willingness cannot be translated into physical action.
Any structural or metabolic disease process that affects the brain diffusely and the frontal lobe in particular can produce apraxia. Selective apraxias exhibit loss of one or more specific movements. The disorder is classified into several types:
The Dominant Hemisphere
Often overlooked is whether the patient is right-handed, left-handed, or ambidextrous. This fact is important in analyzing the patient's complaints because
(2) the dominant hemisphere controls language functions, and
(3) there are many functions that do not appear to be equally represented at a cortical level.
The nondominant (minor) hemisphere plays its major role in nonverbal, geometric, spatial, visual, pattern, and creative or other synthetic functions. Lesions of the corpus callosom will often reveal themselves in dysfunction of the nondominant hemisphere such as illegible writing, constructional apraxia, and the inability to match left-sided tasks easily conducted by the right side.
Heilman and associates list six conditions that can originate from a nondominant hemisphere deficit alone:
(2) inability to get dressed (dressing apraxia),
(3) inability to perceive visual depth,
(4) inability to produce musical sounds (amusia),
(5) inability to recognize familiar faces (prosopagnosia), and
(6) spatial disorientation when verbal clues are absent.
(2) agraphia, the inability to write;
(3) finger agnosia, the inability to count fingers; and
(4) right left confusion, the inability to tell right side from left.
Cranial Entrapment Syndromes
Magoun, Upledger, DeJarnette, and others believe that slight cranial distortions and/or the loss of minute cranial bone mobility may readily lead to CNS dysfunction with expression at the terminals of involved pathways. They attribute many olfactory, optic, oculomotor, trochlear, and other cranial nerve disorders to tension or pressure on these nerves as the result of various direct or indirect effects from the bony components of the skull.
Some Current Subtle Energy Research
How subtle energies affect consciousness and the healing process has long been a mystery though many theories have been formulated. At one time, even speculations of the existence of such energies were placed in the realm of fantasy by most scientists; if they did exist, it was thought that they would be so minute in force that they would have no effect. During the last 2 decades, however, some important advances have been made by the scientific community in trying to determine how the mild stimuli of acupuncture can control pain via the release of endogenous opiate-like substances from CNS centers by directing or releasing such minute foci of energy. These studies also may bring forth new light on the role of subtle vertebral-related neurologic foci in disease processes.
From a viewpoint different from that of many of his medical colleagues, Pelletier, an associate clinical professor at the University of California School of Medicine, has focused attention on developing both a theoretical and a clinical model on how subtle energies (eg, those theorized to be used in acupuncture, homeopathy, and visualization) work in the healing process.
Studies focused on the effects of extremely diluted remedies has seen homeopathic concepts revisited in light of these new findings. Benson, director of the Division of Behavioral Medicine at Beth Israel Hospital at Harvard University is investigating the ancient Chinese practice of chi-gong, an ancient technique for eliciting the relaxation response associated with the alpha state. A joint United States Chinese project is studying the usefulness of chi-gong and herbs in a number of neurologically related diseases.
Zimmerman has been conducting experiments at the University of Colorado on the interaction of living organisms in exterior and interior electromagnetic fields. A biomagnetic imaging system is being developed that will, hopefully, project the electromagnetic field of a living organism as a video image to allow investigation of the biomagnetic correlates of subtle energy healing intervention such as used in therapeutic touch and acupuncture.
At the School of Medicine of the University of North Carolina, Corless bases her project on the proposition that, if emotions are salient to disease onset and progression, they also may be significant to the resolution of disease. A prominent feature of her investigation is the inclusion of drawings, relaxation, mental imagery, and therapeutic touch.
THE POSTERIOR FOSSA LEVEL
The posterior fossa is protected and supported by the temporal bones anteriorly and laterally and by the occipital bones at the base of the skull. The structures within this area serve as the central cerebrospinal level for ascending and descending tracts traveling to and from the supratentorial structures, the brain stem cranial nerve nuclei, the cerebellum, and the spinal cord.
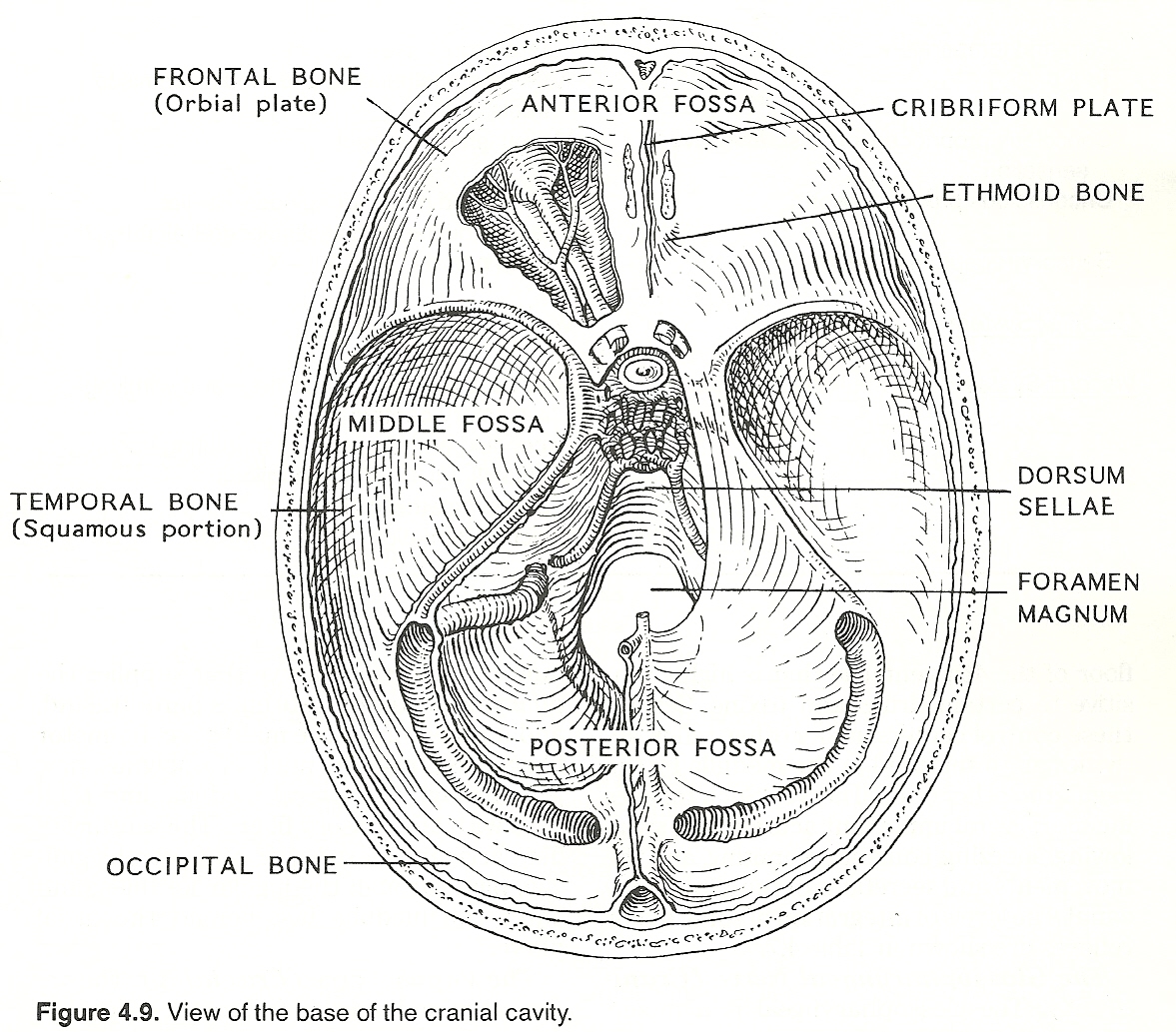
The posterior fossa area is also closely related to the aqueduct of Sylvius (midbrain), 4th ventricle (pons, medulla), and foramina of Megendie and Luschka of the CSF system. It is unique to lesions at the posterior level that unilateral brain stem lesions produce disorders of the ipsilateral side of the face and the contralateral side of the body.

The medulla (spinal bulb) serves as an organ of conduction for the passage of impulses between the spinal cord and the brain. It is continuous with the spinal cord, which, on passing into the cranial cavity through the foramen magnum, widen into a pyramidal shaped mass that extends to the lower aspect of the pons. Externally, the medulla resembles the upper part of the spinal cord, but its internal structure differs considerably.
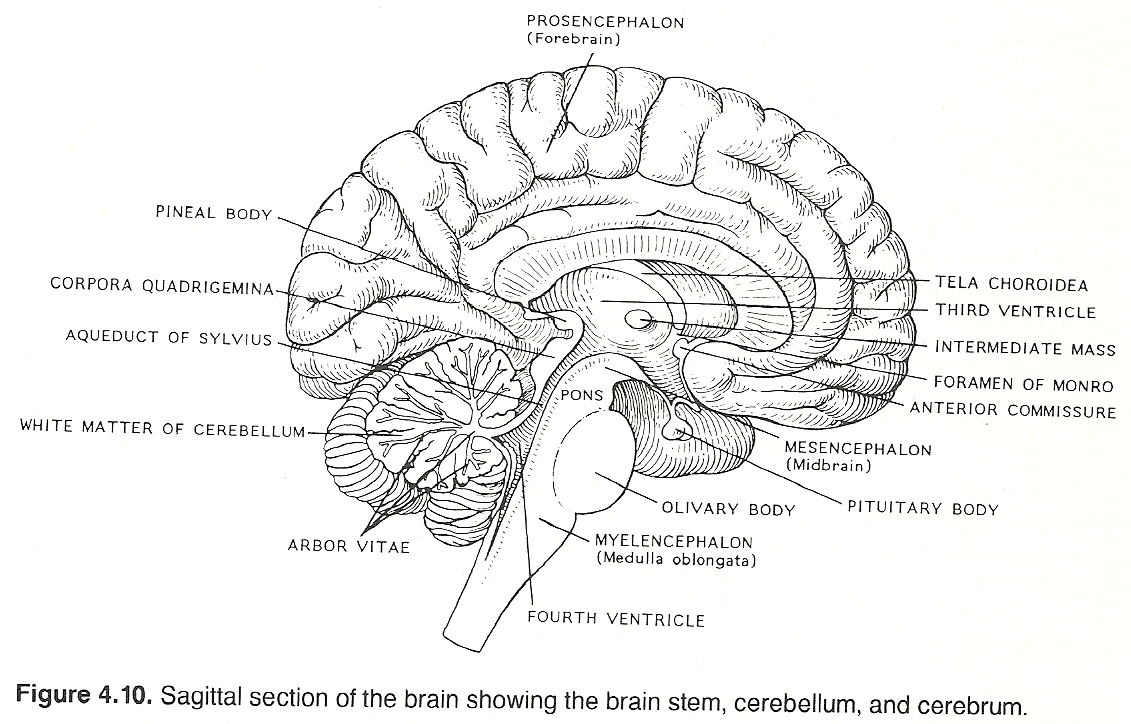
The nerve cells of the medulla are grouped into numerous nuclei, some of which are centers from which cranial nerves arise. The last four cranial nerves (IX XII) originate from nuclei within the medulla. The motor fibers of the glossopharyngeal and vagus nerves and the cranial portion of the accessory nerve arise from the nucleus ambiguus. The hypoglossal nerve arises in the hypoglossal nucleus. From these and other fibers of the medulla, collaterals contribute to the reticular formation.
Some nuclei are relay stations of sensory tracts to the brain such as the nucleus gracilis and nucleus cuneatus, and some serve as centers for body function control such as the cardiac, vasotonic (vasopressor and vasodilator), respiratory (inhalation and exhalation), and emetic centers. There is also a chemoreceptor trigger zone at the floor of the 4th ventricle that is highly sensitive to certain drugs and toxins.
Besides these control centers, many other reflex activities are integrated in the medulla by the vagus and other cranial nerves. Such reflexes include several important acts such as swallowing, sneezing, coughing, vomiting, and the movements and secretions of the alimentary canal.
The levels of integration of some major reflexes are shown in Table 4.7.

The Vagus Nerve (Cranial X). The vagus is a mixed sensory and motor nerve that supplies the pharynx, larynx, esophagus, stomach, heart, lungs, and abdominal viscera, and serves as a pathway for multitudes of afferent and efferent reflex activities. Pharyngoesophageal swallowing, hunger, thirst, cardiac and respiratory rate, peristalsis, and the secretion of digestive juices are just a few examples. Vagal afferents arise from the jugular and nodose ganglia, located below the jugular foramen, pass through the jugular foramen, and enter the medulla just behind the glossopharyngeal nerve. Vagal efferents pass from the medulla and join the course of the sensory fibers.
The Spinal Accessory Nerve (Cranial XI). The multirooted accessory is a motor nerve to the sternocleidomastoideus and trapezius muscles. It differs only from a typical spinal motor nerve by its close connections with the vestibular system and other cranial nerves (eg, orientation of the eyes, nose, and ears toward environmental stimuli). The nerve's rootlets arise behind that of the vagus on the lateral surface of the medulla and upper cervical cord, and they exit the cranial vault via the jugular foramen.
The Hypoglossal Nerve (Cranial XII). The hypoglossal is a motor nerve to the intrinsic muscles of the tongue, thus involved in speaking, eating, and participating in the initial act of swallowing. Like the spinal accessory, the hypoglossal originates from several filaments. These filaments arise on the anterolateral sulcus of the medulla, quickly fuse into one nerve, and exit the posterior fossa of the cranium via the hypoglossal canal.
Basic Functions
Respiratory Center. These bilateral groups of cells in the medulla have an inherent rhythmic activity similar to that of the cardiac center. These centers are especially sensitive to reflexes directed from the cranial and spinal nerves and all pathways between the cerebrum and medulla. Thus, almost any adequate sensory reflex impulses are likely to affect respiratory rate in an excitatory (pressor) or inhibitory (depressor) manner, depending upon the source and effect of the stimuli. Via collateral connections, neurons conveying these impulses affect vasculature tone, heart rate, and cardiac output.
Vasoconstrictor Center. From this bilateral group of cells in the medulla, fibers descend through the spinal cord, and at various levels they form synapses with spinal neurons in the anterior horns of the gray matter, which, in turn, project as preganglionic vasoconstrictor fibers that end in the sympathetic ganglia. Postganglionic fibers then extend to supply most of the peripheral vasculature.
The vasoconstrictor center is normally in a state of continuous activity so that vasculature tone is constant, and different parts of the body are supplied by different paths (eg, skin, intestines). Its activity can be increased reflexly through many types of sensory stimulation to produce vasoconstriction and hypertension (pressor effects), or its activity can be decreased to produce vasodilation and lowered blood pressure (depressor effects).

Medulla Dysfunction
All afferent and efferent tracts of the spinal cord are represented cephally in the medulla, and these tracts either end or decussate at this level. Thus, an isolated medulla lesion may exhibit motor or sensory dysfunction (somatic and/or autonomic) to a degree determined by the site and extent of the destructive process. Lesions of the upper central area of the medulla may involve vital nuclei in this area and produce a large variety of features. Lesions of the posterolateral area produce Wallenberg's syndrome; those of the anterior portion, hypoglossal hemiplegia alternans.
The glossopharyngeal, vagus, spinal accessory, or hypoglossal nerves may be damaged in the medulla or anywhere along their course to the periphery.
The Vagus Nerve. Partial destructive lesions of the cranial vagus exhibit as vagal, recurrent laryngeal, or superior laryngeal paralysis; hysterical aphonia (laryngospasm); or superior laryngeal neuralgia. Caudally, irritative lesions manifest as visceral hyperparasympathicotonia and inhibited lesions as hypersympathicotonia.
In lesions involving the distribution to the throat, symptoms include loss or weakness of voice, chronic cough, dysphonia, dysphagia, esophageal spasm, pharyngolaryngeal pain, salivation abnormalities, and possibly dyspnea and external ear pain. Overt signs include an inhibited gag reflex, an altered position of the vocal cords, bradycardia or tachycardia, and a dilated stomach.
The Spinal Accessory Nerve. Depending on the branch affected, irritative lesions manifest as trapezius and sternocleidomastoideus spasm; inhibiting lesions, as muscle weakness. These conditions can readily be appraised by noting the patient's ability to shrug the shoulders and rotate the head against resistance. In subtle cases, EMG studies may be necessary.
The Hypoglossal Nerve. Injury to the hypoglossal nerve frequently results in paralysis (flaccid or spastic), ataxic tongue, and dysarthria. The patient will complain of eating and speech difficulties. Examination will reveal weak tongue strength when the tongue is pushed to the side of the cheek against external resistance and possibly an atrophic tongue. In addition, lesions of the hypoglossal nerve are frequently combined with other cranial nerve involvement. See Table 4.9.
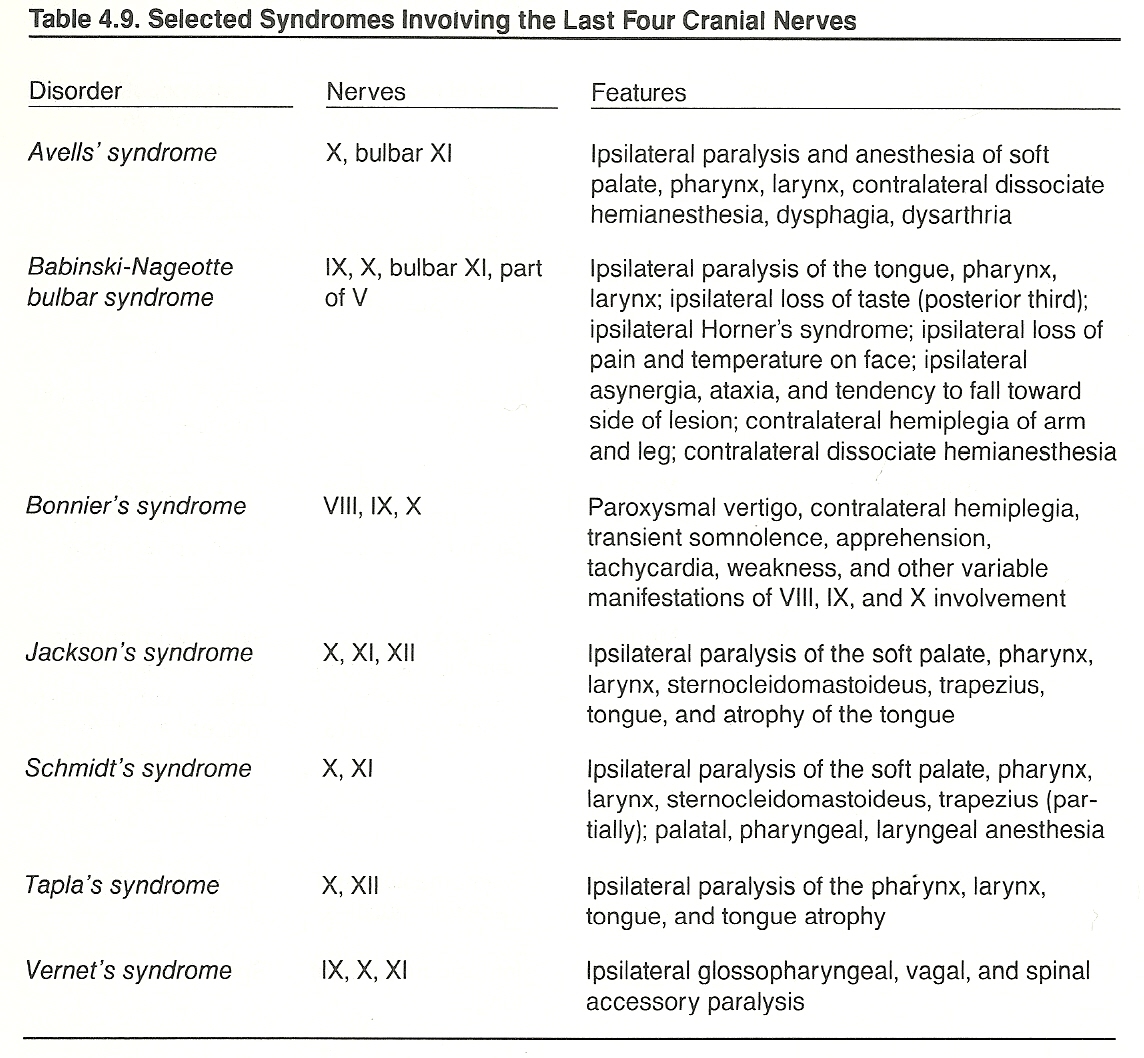
The pons (varoli) serves as a bridge between the two halves of the cerebellum and between the medulla and the cerebrum. It is located in front of the cerebellum between the midbrain and the medulla oblongata and consists of interlaced transverse and longitudinal white nerve fibers intermixed with gray matter. The transverse fibers are those derived from the middle peduncles of the cerebellum and serve to join its two halves. The longitudinal fibers connect the medulla with the cerebrum. Within the pons are the nuclei of all or a part of the fibers of cranial nerves V—VIII.
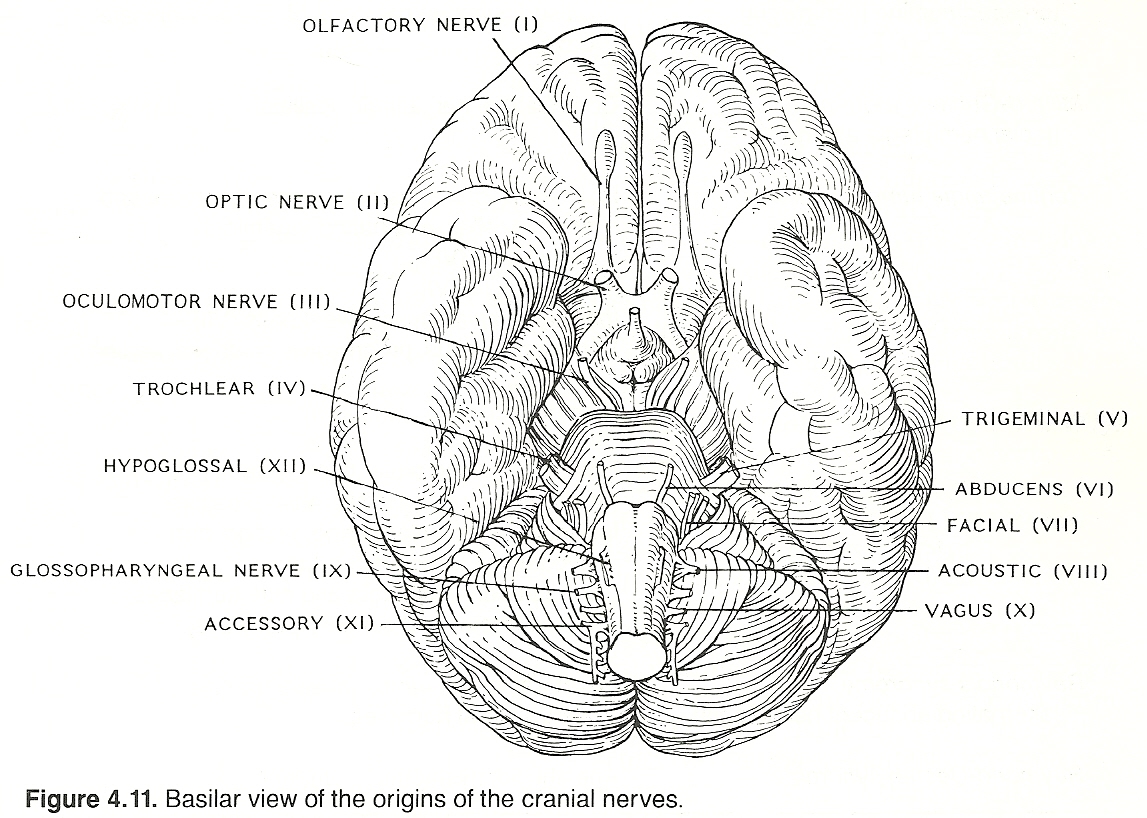
Fibers of the maxillary branch exit via the foramen rotundum, and the ophthalmic branch leaves the cranial vault, via the superior orbital fissure, along with the oculomotor and trochlear nerves. Sensory fibers of the mandibular branch are joined by the motor fibers of the nerve, which leave the pons anteromedially to the sensory fibers and exit the skull via the foramen ovale.
The Abducens Nerve (Cranial VI). The abducens nerve supplies motor fibers to the lateral rectus muscle of the eye. When activated, the eyeball rolls laterally. It arises from the anterior surface of the brain stem at the groove that externally separates the pons and medulla. It then passes through the cavernous sinus and exits the skull via the superior orbital fissure, along with cranial III, IV, and V (ophthalmic branch) nerves.
The Facial Nerve (Cranial VII). The facial nerve is a mixed sensory and motor nerve that supplies the muscles of facial expression; the muscles necessary for hyoid elevation; the tongue's anterior taste buds; and the lacrimal, nasal, palatal, sublingual, and submaxillary glands. The sensory root originates from the cells of the geniculate ganglion, passes through the internal acoustic meatus and end in the medulla. The motor root projects from the posterior pons and exits the skull via the internal auditory meatus.
The Acoustic Nerve (Cranial VIII). The two-part acoustic nerve is the sensory nerve of hearing (middle ear) and balance (inner ear). The cochlear branch originates from the spiral ganglion, and the vestibular division arises from the vestibular ganglion (ganglion of Scarpa). The latter is in the posterior aspect of the internal auditory apparatus. The acoustic nerve enters the cranium via the internal acoustic meatus and enters the penetrates the brain stem just behind the posterior edge of the middle cerebellar peduncle.
Pontine Dysfunction
Lesions of the lateral pons (eg, pontocerebellar angle tumor) result in cranial nerve V—VII, IX X, and XII dysfunction; alternating cranial V hemiplegia; ipsilateral ataxia, vertigo, hypotonia; contralateral hemiplegia, hemianesthesia; and general signs of increased intracranial pressure. Lesions of the anteroinferior pons frequently produce alternating abducent hemiplegia, Foville's syndrome, or Millard-Gubler's syndrome.
The effects of common lesions in the pons are shown in Table 4.10.
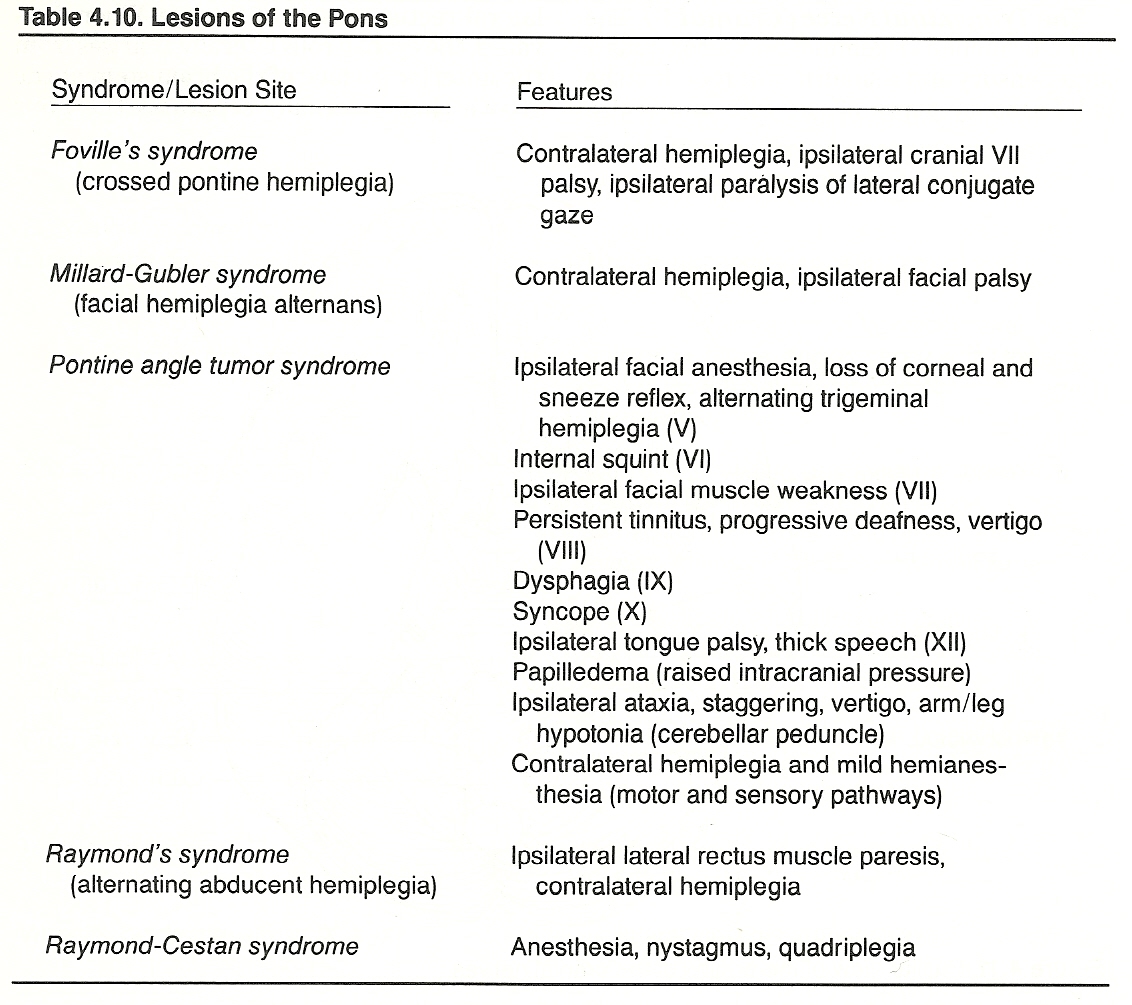
The Abducens Nerve. As the abducens is the motor nerve for ocular abduction, lesions produce paralysis of this function: medial in inhibiting lesions, lateral in irritating lesions. Both types will have associated diplopia.
The Facial Nerve. Lesions of the facial nerve most frequently exhibit as Bell's palsy, the nuclear or supranuclear types of facial palsy, geniculate neuralgia, facial spasm (paroxysmal facial hyperkinesia), bilateral facial palsy (facial diplegia), striatum lesions, and crocodile tears syndrome (paroxysmal lacrimation). Symptoms of facial nerve involvement include pain and facial motor abnormalities. The overt signs of a facial nerve lesion include the inability to smile or whistle, hyperlacrimation, facial twitching or paralysis (usually unilateral), facial asymmetry, and impaired taste on the anterior 2/3rds of the tongue.
Cochlear Division of the Acoustic Nerve. The common symptoms of a lesion of the cochlear branch include deafness, tinnitus, hearing scotomas, and auditory hallucinations. The overt signs are diminished hearing acuity and sensory aphasia.
Vestibular Division of the Acoustic Nerve. Symptoms of a lesion of the vestibular branch include spatial disorientation, vertigo, and nausea. The major signs of vestibular nerve involvement are nystagmus, diaphoresis, hypotension, tachycardia, and vomiting.
The Midbrain
The midbrain (mesencephalon) is a short, constricted portion that connects the pons and cerebellum with the hemispheres of the cerebrum.
The Trochlear Nerve (Cranial IV). The trochlear nerve provides the major motor supply to the superior oblique muscle of the eye. When stimulated, the eyeball rolls down and medial. The nerve arises from the posterior surface of the brain stem, curves anteriorly between the posterior cerebral and superorcerebellar arteries, and continues forward (between the oculomotor and ophthalmic branch of the trigeminal) in the lateral wall of the cavernous sinus. It enters the orbit of the eye, along with the oculomotor nerve, via the superior orbital fissure.
Midbrain Dysfunction
The effects of common lesions in the midbrain are shown in Table 4.11.
The Trochlear Nerve. Inhibiting syndromes affecting the trochlear nerve prevent the patient from looking down and nasally.
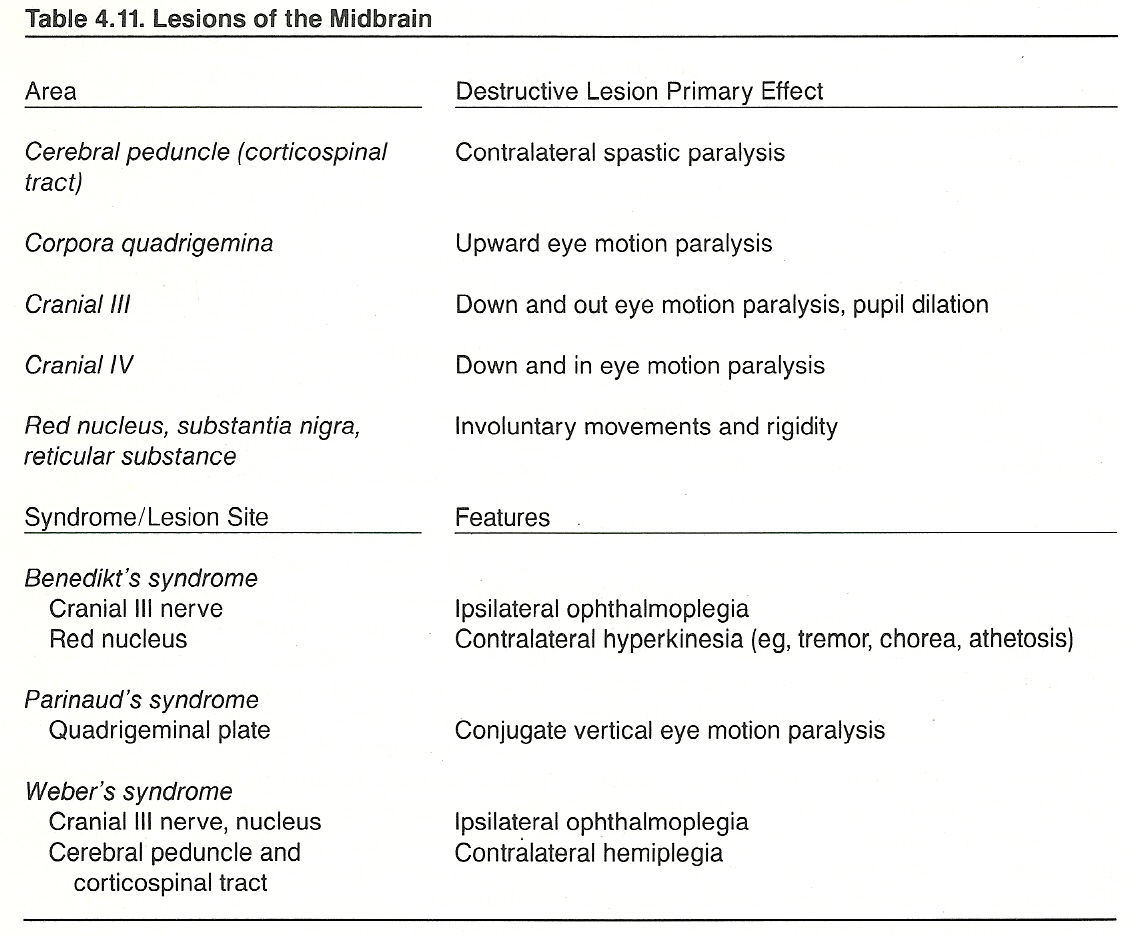
The cerebellum develops embryologically as an outgrowth from the primary centers of the semicircular canals in the medulla. It is below the posterior portion of the cerebrum, from which it is separated by the tentorium cerebelli (a fold of dura mater), and behind the pons and upper aspect of the medulla. The oval cerebellum appears constricted in its center (vermis), flattened from above downward, and has lateral expanded portions (the hemispheres).
Structure
The cerebellum is divided into three lobes that have specific functions. The large posterior lobe is responsible for coordinated movements of the extremities, the flocculonodular lobe directs equilibrium and balance, and the anterior lobe is concerned with gait and posture.
The gray matter surface of the cerebellum is not convoluted; rather, it is transversed by numerous furrows (sulci). The gray matter contains cells from which fibers pass to form synapses in other areas of the brain and cells with which fibers entering the cerebellum from other parts of the brain form synaptic complexes. The internal structure of the cerebellum contains various important centers such as the dentate, emboliform, fastigial, globose, and interpositus nuclei.
The myelinated tracts of the cerebellum are connected with the cerebrum by the superior peduncles, with the pons by the middle peduncles, and with the medulla oblongata by the inferior peduncles. Afferent impulses from the motor centers of the cerebrum, the semicircular canals of the inner ear, the olivary body, and somatic muscles enter the cerebellum via the three peduncles (Table 4.12). Efferent impulses are transmitted to the motor centers in the cerebrum, to several cranial nerve nuclei, and down the brain stem and spinal cord to connect with somatic muscles. Synaptic connections occur at many levels.
Function
Activities of the cerebellum do not enter normal consciousness. The cerebellum receives sensory impulses from the cerebral cortex, semicircular canals, body wall, and somatic muscles. It sends impulses into all somatic motor centers and helps to maintain equilibrium and the tone of voluntary muscles.
Dysfunction
Cerebellar lesions result in muscle weakness, loss of tone, and the inability to direct precise movements of skeletal muscle. There is usually difficulty in walking due to inability to control the muscles of the legs or difficulty in talking due to lack of coordination of the tongue and jaw muscles. The area of the body affected is determined by the location and extent of the damage. Only parts of the body on the ipsilateral side of the injury to the cerebellum are involved. If the damage is bilateral, then the lack of muscle tone and coordination may be so great that the patient is physically helpless.
Clinical Considerations
See Chapter 8 and
Chapter 9.
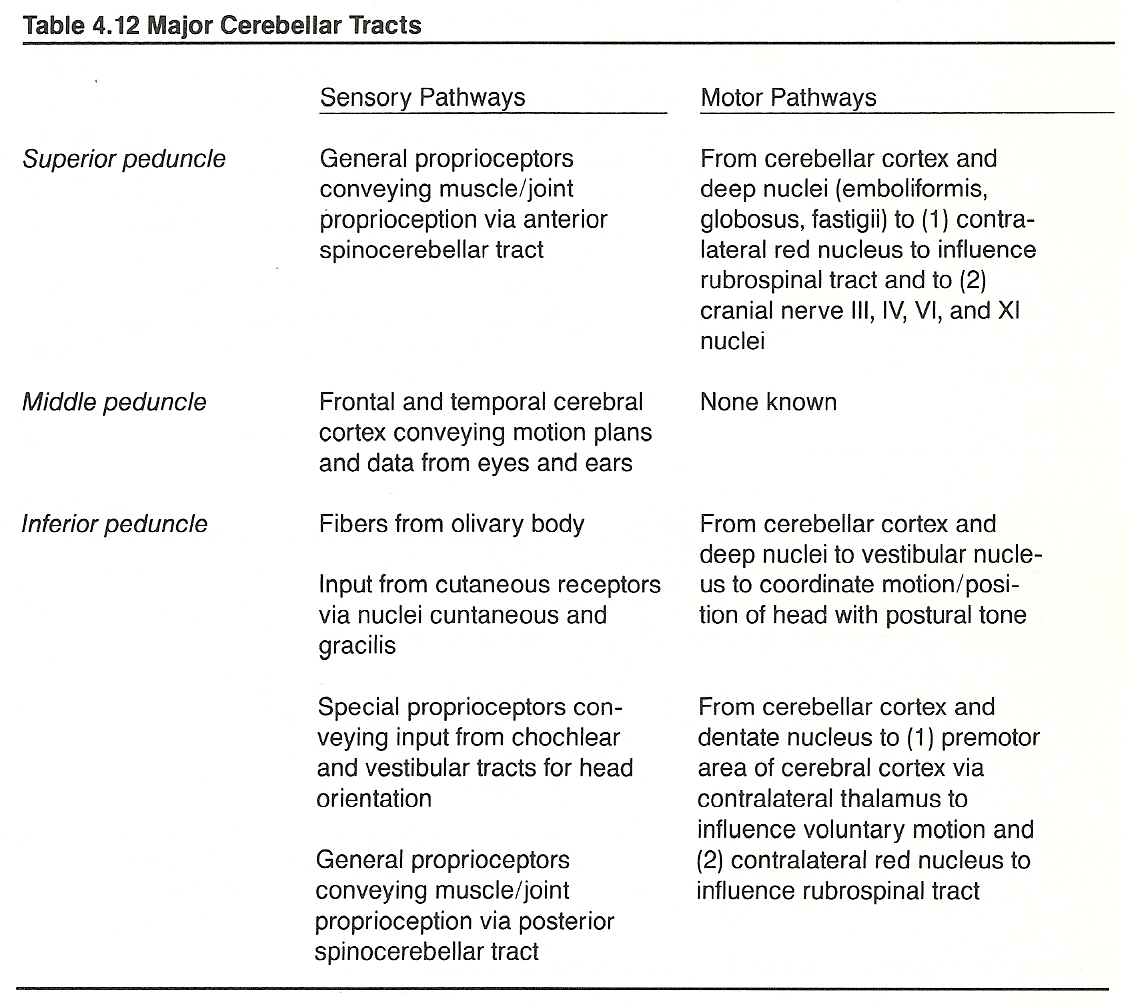
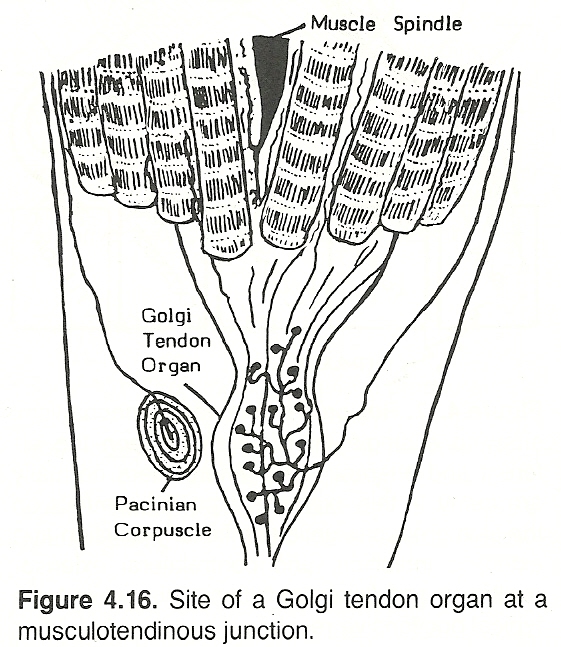
Sensory input from Golgi tendon organs, pain receptors, and spindle flower-spray terminals to the spinal cord are transmitted to motor neurons via one or several interneurons and synapses that may have either an excitatory or inhibitory function. Through these multisynaptic routes, afferent impulses can be transmitted ipsilaterally, contralaterally, and inferiorly or superiorly to adjacent segments to excite accessory muscle groups or inhibit antagonists. Thus, each motor neuron is subject to numerous segmental influences as well as influences from higher centers transmitted via the descending pathways and long loops.
Nocifensive Reflexes
The primitive polysynaptic nociceptive reflexes are sometimes called withdrawal, cutaneous, or flexion reflexes. The receptors of such reflexes originate from bare nerve endings within tissue (usually superficial) and are conveyed over small myelinated and unmyelinated
(2) muscle Group III and C afferents to multisynaptic relays of the anterior motor neuronal pool, which has a residual after-discharge effect that outlasts the stimulus.
Abnormal nocifensive reflexes are diagnostic of CNS lesions. Babinski's plantar reflex in corticospinal tract disease is a classic example.
Sustaining Mechanisms
Reflexes such as stretch reflexes and inverse myotatic reflexes are self-limiting mechanisms whereby sensory impulses from muscle are transmitted to the same muscle via the afferent cord efferent reflex cycle. This type of activity, however, does not explain sustained activity such as seen in voluntary phasic movements. Voluntary activity is initiated and controlled by brain centers. It does not depend upon sensory stimulation, but it does include the segmental reflex centers, especially the stretch reflex circuits. Voluntary action involves loop coactivation of gamma and alpha efferents.
Gamma Neuron Reflexes
Movement can be initiated by activating either alpha or gamma motor fibers. When alpha motor neurons are stimulated, extrafusal muscle fibers contract and the length of the muscle spindles reduces. However, for a muscle spindle to respond appropriately to a change in tension, its length must be adjusted.
This is accomplished by separate innervation (fusimotor system) via the plate and trail endings of gamma motor fibers. When the lower threshold gamma motor neurons are activated, the effect is on intrafusal muscle fibers of the muscle spindles. Gamma neurons receive their input segmentally and supraspinally from the corticospinal, reticulospinal, and propriospinal tracts.
Renshaw Inhibition
Renshaw cells are inhibiting interneuronal cells within Rexed's laminae VII and VIII of the anterior horn of the spinal cord. They project axons that interact with alpha motor neurons that enervate extrafusal muscles. When fast-twitch alpha motor fibers are activated, Renshaw cells become activated, which, in turn, temporarily inhibit the small alpha fibers of synergistic motor neurons (recurrent inhibition), thus acting as a stabilizer to subdue postural (tonic) stretch reflexes during phasic motor activity.
Righting and Placement Reflexes
Stretch reflexes, integrated in the spinal cord and medulla, are just one type of postural reflex; ie, an automatic response to keep the top side of the body or a body part properly positioned (eg, uppermost). Optic righting reflexes attempt to maintain the eyes on a horizontal level and depend on an intact occipital cortex. Labyrinthine righting reflexes attempt to maintain orientation of the head in space and require an intact midbrain.
Cervical righting reflexes arising in the neck attempt to maintain the body in proper relationship with the head and are integrated in the medulla. There are two types of body righting reflexes:
(2) body righting arising from surface receptors that act on the body in an attempt to maintain orientation in space. Both these types of body righting reflexes require an intact midbrain.
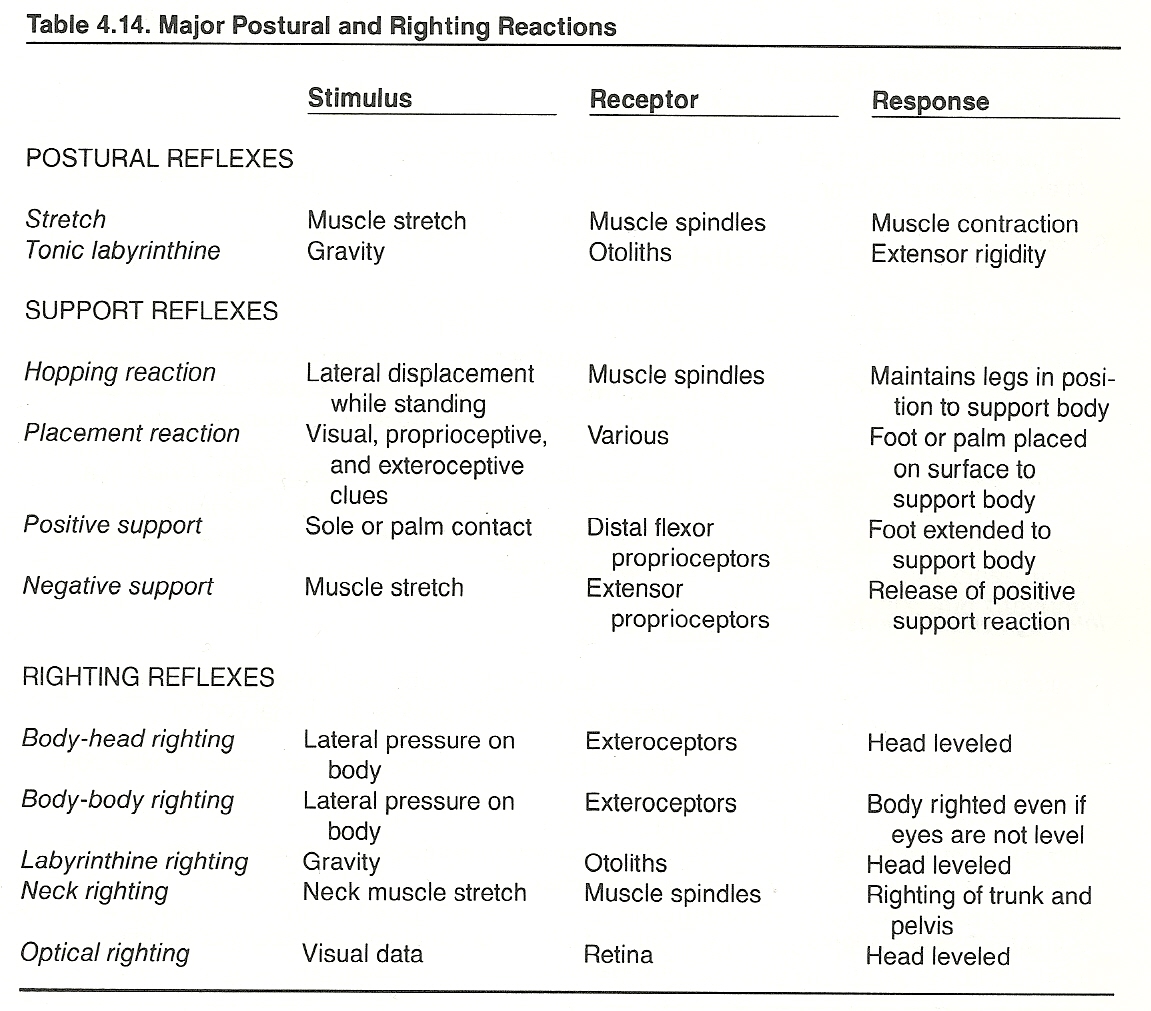
Diagnostic Considerations
Part III of this book is directed almost exclusively to the study of clinical disorders originating at the spinal level. However, a few points are worthy of mention here.
At almost every level, the spinal level contains representatives of the CSF, motor, sensory, vascular, and visceral systems; thus, lesions will have widespread implications. Because the spinal level is the intermediary between the longitudinal and horizontal peripheral systems below the foramen magnum, a lesion at a spinal level may express longitudinal, peripheral, or combined manifestations.
Diseases at the spinal level can be classified into focal segmental (eg, IVF syndromes), focal intersegmental (eg, involving both segmental and longitudinal fibers), or diffuse (eg, meningitis, degenerative disorders).
Typical signs are shown in Table 4.15.

Injuries to the lumbar cord or its tail occur from vertebral fractures, dislocations, or penetrating wounds in severe accidents. In rare instances, the cord may be damaged from violent falls with trunk flexion. The T12—L1 and L5—S1 areas are the common sites of injury, especially those of crushing fractures with cord compression.
Neurologic symptoms develop rapidly, but the lower the injury, the fewer roots will be involved. More common than these rare occurrences are cord traction, concussions, and contusions. Even a trace of sensory abnormality, objective or subjective, should immediately raise the suspicion of injury to the spinal cord or cauda equina.
Contusions of the Spinal Cord. Cord concussion usually complicates cord contusion. If laceration occurs, shock is rapid. Deep reflexes, sensation, and sphincter control are lost. The paralysis is flaccid. Obviously, a prognosis cannot be made until the shock is survived.
Pathologic Traction of the Spinal Cord. A scoliotic lumbar deviation is usually attended by a commensurate vertebral body rotation to the convex side. If this does not occur, it is atypical and most likely pain producing. If the vertebral bodies were not subject to the law of rotation during bending, the spine would have to lengthen during bending and its contents (ie, cord, cauda equina, and their coverings) would be subjected to considerable stretch.
Reflex Phenomena in Spinal Disorders
Sandoz believes that the main reflex phenomena associated with vertebral disarrangements and adjustments pertain to referred pain, the regulation of tonic and phasic activity of the innervated paraspinal musculature, the peripheral modulation of pain perception, and tonic neck reflexes.
THE PERIPHERAL LEVELS
Peripheral nerve histology and end-organs function have been described previously. Thus, mention of these subjects in this section will be brief. To review fiber class, diameter size, conduction velocity, and function,
refer to Table 3.4.
The Peripheral Nerves
Each peripheral nerve has a well-defined course in textbooks. Such descriptions are averages, and it is not uncommon to see this course vary from 1 to 2 inches at various sites in individuals once they have extended well beyond their origins in the CNS. Things are not always as they should be, and to believe otherwise may lead to erroneous conclusions.
As each nerve progresses, it usually branches often, and thick branches divide again and again, until a fine network of minute twigs is formed. This can be likened in many respects with the design of the tree, with the trunk of the tree representing the spinal column. Within his 1925 text, Riley stated: "So completely are nerves distributed to parts of the body that were everything but the nerves removed, they alone would preserve the shape of the body in all its outlines."
The Peripheral Plexuses
A nerve plexus is a complex network of peripheral axons. There are three large somatic plexuses: the cervical, the brachial, and the sacral (lumbosacral). Refer to Table 3.12 for the neurologic signs exhibited in the cervical and lumbosacral radiculopathies. As weakness is a common sign in impairment of somatic effectors, testing distal muscle strength will often aid in localization (Table 4.16).
The Posterior Rami
Shortly after the junction of the anterior and posterior roots, a typical spinal nerve divides into three divisions:
(2) a posterior primary division, and
(3) an anterior primary division.
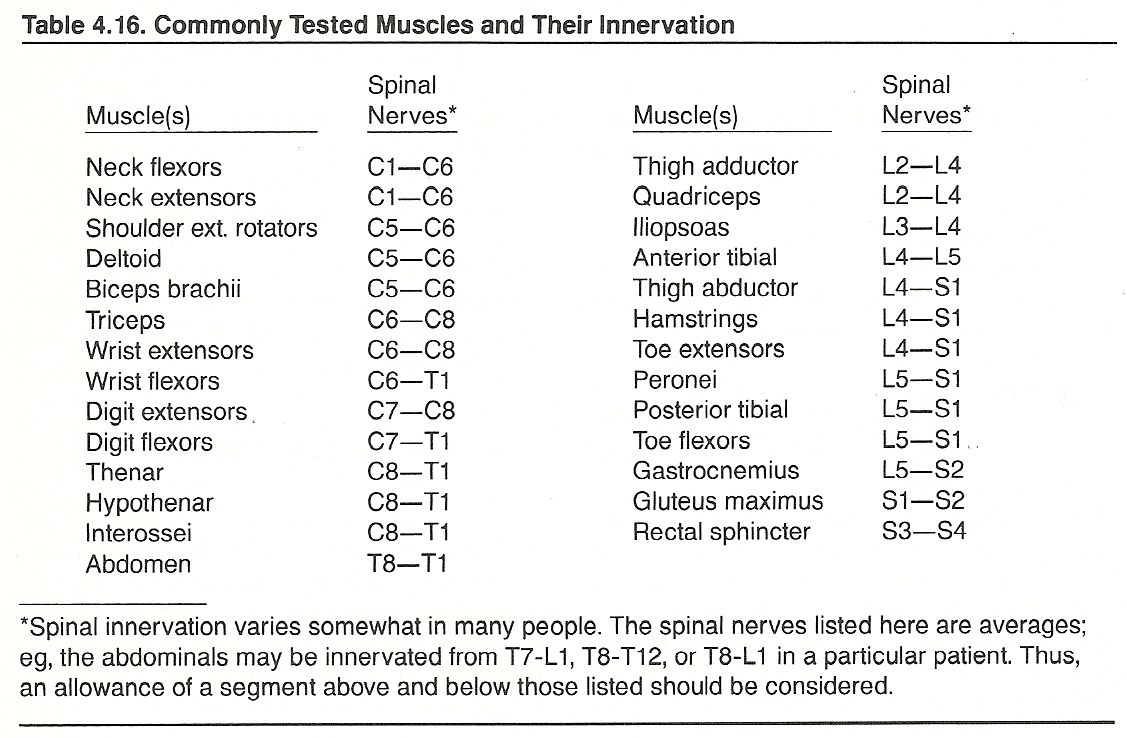
Each cylindrical muscle fiber is covered with a specialized layer of sarcolemma that covers a peripheral longitudinal layer of nuclei and cylindrical myofibrils. Sacrolemma insulates each fiber from adjacent fibers so that excitation of one fiber does not influence its neighbors.
The basic functional unit of muscle is the sarcomere, many of which are connected end to end to form a single myofibril. About a thousand myofibrils, laying side by side, are contained within each muscle cell. Dispersed in a regular pattern among myofibrils is the sacroplasmic reticulum, which is an extensive membrane system of minute tubes; viz, the T-tubule system. This system is the major site of calcium sequestration within muscle tissue.
The tunnels of the T-tubule system, unique to muscle cells, extend completely through the cell and are continuous with the extracellular space at both ends. The fluid within them is primarily extracellular fluid that is thought to be altered by the interaction of its biochemical components with the extremely thin walls of the narrow tubules. Each tubule within a muscle cell is closely associated with several enlargements of the sacroplasmic reticulum (cisterns). Some unknown signal passes from the tubules to the cisterns during excitation during the contraction process.
Within each muscle fiber, there are a thousand or more longitudinal myofibrils arranged in bundles that are about 80% of fiber volume. Each myofibril contains many white I bands and many dark, short, thicker A bands, and each of these isotropic and anisotropic bands contains specialized zones. It is the stacking of myofibrils in register that gives voluntary muscle its microscopic transverse striations.
The thin white bands of myofibrils contain smaller myofilaments composed of actin, tropomyosin, and troponin protein; the thick dark bands consist primarily of myosin protein. An A band contains a central H zone composed solely of myosin filaments that interdigitate with the thin actin filaments on either side. The midpoints of the myosin filaments are secured to one another transversely by connections whose line of cross links is termed the M line at the middle of the H zone.
Gross muscle shortening should not be confused with filament shortening. During excitation, the contractile elements do shorten; however, whether the gross muscle shortens or not depends on the relation of the internal muscle force developed to the load. At the ultramicroscopic level, it is believed that the contractile elements (actin and myosin) also slide into an overlapping interdigitated arrangement upon excitation.
The Neuromuscular Junction
The neuromuscular junction can be considered a specialized type of neuroeffector synapse. The myelin sheath of the terminal axon narrows and then ceases just prior to the motor end-plate. At the site of the end-plates, capillary anastomoses are especially well developed to maintain good nutrition.
Nerve fibers enter the endomysium but do not penetrate the sarcolemma; their terminal branches appose the surface of the muscle fiber. This site is a specialized area of axolemma and sarcolemma where the subneural apparatus is folded into clefts and the sarcolemma is abundant with mitochondria. It is a specialized area that serves as a minute structural barrier where the nerve impulse must be transmitted chemically.
Axon terminals contain mitochondria and numerous vesicles that store the chemical bridge acetylcholine which allows the nerve impulse to be transmitted across the synapse. It is thought that the transmitter substance changes sarcolemma permeability to allow an influx of sodium ions that generate the electric potential to release the calcium necessary in turn for the acetylcholine release that allows the interaction of myosin and actin filaments to produce muscle contraction.
The enzyme cholinesterase has a dual function. It can activate nerve sheath acetylcholine into its active form or degrade it into acetic acid and choline. Thus, a lack of cholinesterase can either produce abnormal or subnormal function. This often manifests as jerking skeletal muscles just before or during sleep.
This sign is thought to be due to a lack of cholinesterase that prevents residual acetylcholine from being properly inactivated. A complaint of frequently dropping objects involuntarily also may represent a shortage of cholinesterase failing to inactivate acetylcholine. Such signs may be a clue to otherwise subclinical riboflavin and/or niacin deficiency that may be expressed in neuromuscular biomechanical mechanisms.
Impulse Response
Nerves containing both motor and sensory fibers pierce muscle fascia at the site of the primary arterial branch (the neurovascular hila). These nerves then fan out to supply both motor and sensory nerve fibers to each muscle spindle.
With the arrival of nerve impulses, usually below 50/sec, all muscle fibers of a motor unit contract synchronously and contract to their maximum (all-or-none law). When several motor units are stimulated by asynchronous firings, the attached fibers contract, and voluntary body movement occurs. However, because motor units fatigue after several stimulations, other units must be called into play to allow for smooth movement. While it appears during normal muscle contraction that all muscle fibers are in a smooth continuous muscle contraction, they are not. This appearance results from the sum of a series of small groups of fibers contracting at the same moment.
All muscle fibers of a motor unit do not lie in an adjacent position. They are spread throughout the muscle belly so that excitation will produce a weak contraction in a broad area of the muscle, rather than a strong contraction at an isolated area. This allows delicate manipulations.
Muscle Spindles and Their Role in Motor Reflexes
It has been previously described that the alpha motor neuron must fire if a skeletal muscle is to contract; thus its designation as the "final common pathway" for motion. The major sources of input are the muscles spindles and the higher CNS centers related to voluntary movement. Muscle spindles also provide signals to the CNS about muscle stretch via Ia fibers and about steady muscle tension over group II fibers.
As described earlier, each muscle spindle contains a few muscle fibers whose purpose is not to help in contracting the whole muscle but to control the sensitivity of the muscle spindle to stretch. Spindle fibers are controlled by gamma motor neurons, while other muscle fibers are activated separately by alpha motor neurons. DeBacher has found that the more contracted muscle spindles become, the more sensitive they are to stretch stimuli.
If a tendon of an agonist muscle is tapped, the muscle and the spindle within it are lengthened and this excites the primary spindle receptors (the annulospiral endings) at the ends of the Ia fibers. This causes volleys of action potentials to course along many Ia fibers to the spinal cord wherein they excite many alpha motor neurons, whose action potentials, in turn, cause the muscle to contract. This concise explanation of these mechanisms by Prichard goes on to note that the synapse transmitter between the Ia terminals and the motor neuron pickup is unknown but thought to be of an electrical rather than chemical nature.
When the Ia volley stimulates motor neurons regulating its own muscle, it retards those regulating the antagonists through a disynaptic pathway. Again, the mode of impulse transfer to the inhibitory interneuron is unknown, but it is understood that the interneuron releases glycine which produces an inhibitory postsynaptic potential (IPSP) on the antagonist motor neuron.
The role of muscle spindles is to provide data to the CNS about muscle stretching rates. The spindles do this through the action potentials conveyed within the train of volleys conducted by the Ia fibers. Using this information plus signals concerned with limb position, the CNS computes the general position of the limb at some near future time. This calculated guess is necessary because without the element of anticipation, the time required for conduction from sensory receptors to cord, central processing, and conduction to muscle would be too long to regulate rapid motion.
The gamma (fusiform) motor neurons regulate the sensitivity of the spindle system by directing the shortening of the modified intrafusal muscle fibers. These fibers are in series with the central-receptor aspects of the spindles. As with alpha motor neurons, gamma fibers receive signals of many types and from various sources, including each other and the alpha motor neurons. It should be pointed out, however, that this complex segmental apparatus is only the lowest aspect of a highly integrated CNS system for central motor control.
The Contraction Process
When a decision is made at the supratentorial level to move a part (ie, contract the muscle fibers of specific motor units), a chain of command is invoked that has an action potential which is transmitted down the spinal cord and then peripherally to the appropriate neuromuscular junctions. From here, the action potential spreads from the end-plate area of the muscle cell along its plasma membrane, and acts upon the muscle cells to trigger the contraction process.
The action-potential process for muscle fibers is similar to that described for nerve fibers. The mechanism is initiated on the electrically excitable membrane surrounding the chemosensitive but electrically inexcitable end-plate by currents flowing through end-plate channels caused to conduct by acetylcholine.
When a nerve impulse reaches the neuromuscular junction, a transmitter is released that diffuses through the barrier to excite the muscle fiber. The energy potential enters the muscle fiber, via the minute T tubules, then travels to the A and I band junctions where it triggers the release of calcium, which initiates the process of fiber activity. Myofibril contractile elements tense as the action potential travels across the sarcolemma. The rate is 1—3 m/sec.
During rest, the thick myosin myofilaments only slightly overlap the thin actin myofilaments. During excitation, the A and I bands are thought to slide increasingly into an overlapping position to a degree proportional to the chemical interactions between filaments. The light bands appear to shorten and finally disappear as the thin filaments slide toward each other between the dark myofilaments. When the action potential ceases, calcium returns to storage, the elastic elements recoil, and the thin I bands draw back to their relaxed state.
Contraction Strength
Muscle power is equal to the load moved times the speed at which the load is moved. The amount of shortening (strength) of a muscle fiber following a stimulus depends on a combination of such complex variables as:
(2) the specific characteristics of the stimulus,
(3) the speed at which the muscle is required to act,
(4) the rate of flow of nerve impulses arriving from each active nerve fiber,
(5) the degree of stretched fiber elongation at the beginning of contraction,
(6) whether the muscle tissue is warm or cool, and
(7) the state of local fatigue and nutrition. Although maximum effort may be applied to a task, biomechanical performance will be either enhanced or diminished by these conditions.
Clinical Considerations
From a clinical standpoint, lesions of peripheral nerves are divided into disorders of nerve proper and disorders of their end organs. This is because impairments of nerves have different characteristics than do disorders of end-organs.
Focal and Diffuse Disorders
Diseases involving peripheral nerves may be either focal (involving one nerve) or diffuse (involving whole areas of peripheral nerves).
Diffuse Disorders. Widespread lesions may be the result of poisoning, severe trauma, or a wide variety of degenerative, vascular, inflammatory, metabolic, endocrine, and neoplastic disorders. Either the myelin, the axons, or both may be involved, but in each instance, the clinical features are similar:
(2) paresthesia, dysesthesia, and hyperalgesia;
(3) absent or diminished reflexes; and
(4) distal muscle weakness leading to faccidness, atrophy, and deformity. Disorders involving primarily the axons feature early distal muscle fibrillation and severe atrophy. Disorders involving primarily the myelin are characterized by little or no early fibrillation and only minor atrophy.
Specific Peripheral Nerve Disease
Diseases involving peripheral nerves usually exhibit combined sensory, motor, and autonomic signs and symptoms. Typical sensory/motor examples include segmental distal weakness and atrophy, absent or diminished sensations, and segmental loss or impairment of superficial and deep reflexes. If autonomic fibers are involved, temperature, perspiration, and trophic abnormalities will manifest. Causalgia also may be reported.
Neuronal Plasticity in the Healing Process: A Brief Review
When axons are injured by either trauma or a disease process, Bishop describes the process as an initial sealing of the cut ends. This is followed by orderly degenerative and phagocytic events in the segments lying distal and proximal to the lesion. Injured axons lose their dendrites and, thus, their synapses.
Electrophysiologic and biochemistry properties are altered to prepare the injured neurons for synthesizing proteins needed for the formation of growth cones and regeneration of the fibers. During this period, the denervated muscle, if one is involved, undergoes dramatic atrophic changes that not only modify the electrical and chemical properties of its membrane but also reduce the diameter and weight of its fibers.
The response to injury of sensory axons depends on the site of the lesion. Cutting the fibers of a posterior root, for example, causes Wallerian degeneration of the primary afferents' central processes. Collateral sprouts from neighboring uncut root fibers are thought to take over the vacant synapses on the 2nd order neurons in the dorsal horn.
Cutting a sensory axon peripherally eliminates the receptive field of the primary afferent initially, but in time, collaterals from neighboring cutaneous afferents provide the 2nd order neurons with a new receptive field. Synaptic reclamation and receptive field reorganization are but two dramatic examples of neuronal plasticity that lead to "rewired" neural circuitry.
Motor Axonal Impairments
A major sign of early motor impairment is fasciculation. Fasciculation (a small twitch under the skin) reflects a low-threshold firing somewhere in the motor unit (commonly in the efferent fiber), that is producing neuronal irritability and possible spontaneous firing. A similar hypersensitivity in a sensory fiber is perceived by the patient as pain if the small fibers are irritated or as a paresthesia (eg, tingling) if the large fibers are involved.
Axoplasmic Disorders. A nerve's axoplasm and its flow are commonly disturbed by laceration, heavy pressure, or other types of severe injury. In some chronic slowly progressing disorders, fibers tend to die distally (axonal atrophy) such as seen from local compression and in various deficiency, genetic, metabolic, and toxic states. Wallerian degeneration follows in both acute and chronic forms.
Note: In situations of local compression (eg, an entrapment syndrome), moderate narrowing of the axons involved results in a slowed conduction speed. Such axons will still transmit impulses but at subnormal rates. The higher frequency impulses are affected most; thus, most noticeable will be impairments in
(1) muscle spindle output,
(2) rapid vibration perception, and
(3) muscle contraction deficiency.
Myelin Disorders. A loss of myelin results in slowed conduction velocity as reflected in loss of reflexes and strong muscle contraction, and impaired vibration and proprioceptive sensations. As demyelinization progresses, action potentials will become completely blocked. Genetic, immunologic, and toxic disorders are the common causes.
Neuromuscular Trophic Impairments
Efferent nerves supplying muscles provide an important factor in muscle nutrition. Because of this, a denervated muscle will undergo membrane changes and atrophy. There are four common forms of this disorder:
In striated muscle trophic impairment, alpha efferents are involved and a spreading hypersensitivity of acetylcholine receptors results that starts adjacent to the end-plates. Fibrillation (rhythmic twitching of single muscle fibers) is an early finding.
In anterior horn cell involvement, one or more descending pathways are involved and hypersensitivity results from local sensory input. Spasticity, hyperreflexia, and clonus are early findings.
In destruction of postganglionic parasympathetic fibers of the oculomotor nerve, hypersensitivity to acetylcholine analogs results. Mydriasis (abnormal pupil dilation) is an early sign.
In destruction of postganglionic sympathetic fibers of the oculomotor nerve, hypersensitivity to epinephrine analogs results. Miosis (abnormal pupil constriction) is an early feature.
Neuromuscular Junction Disease
Neuromuscular junction disease (NMJD) mimics and differs from axonal disorders in several respects. There is no atrophy, reflexes and muscle tone are normal, sensory perception is intact, mental function is normal, and no signs are evident of longitudinal tract involvement. The major features of NMJD are fluctuating weakness upon exertion (usually in proximal or cranial muscles). See Table 4.17.
Peripheral Nerve Pinch, Stretch, and Compression Syndromes
Nerve "pinch" or "stretch" syndromes are especially common in sports. They are also seen after falls and industrial accidents, and the syndrome effects appear throughout the cranium, spine, pelvis, and extremities. Hardly any peripheral nerve is exempt.
A nerve pinch syndrome may be due to direct trauma (contusion and swelling), subluxation, a protruded disc that results in nerve compression, sclerosis, or fracture (callus formation and associated posttraumatic adhesions). Any telescoping, hyperflexion, hyperextension, or hyperrotational force to the spine may result in a nerve pinch syndrome where pain may be local or extended distally. Peripheral nerve pinch syndromes are less common than nerve stretch syndromes but are usually more serious.
A nerve stretch syndrome is commonly associated with sprains, severe lateral cervical flexion with shoulder depression, or dislocations. Nerve fibers may be stretched, partially torn, or ruptured most anywhere in the nervous system from the cord to peripheral nerve terminals.
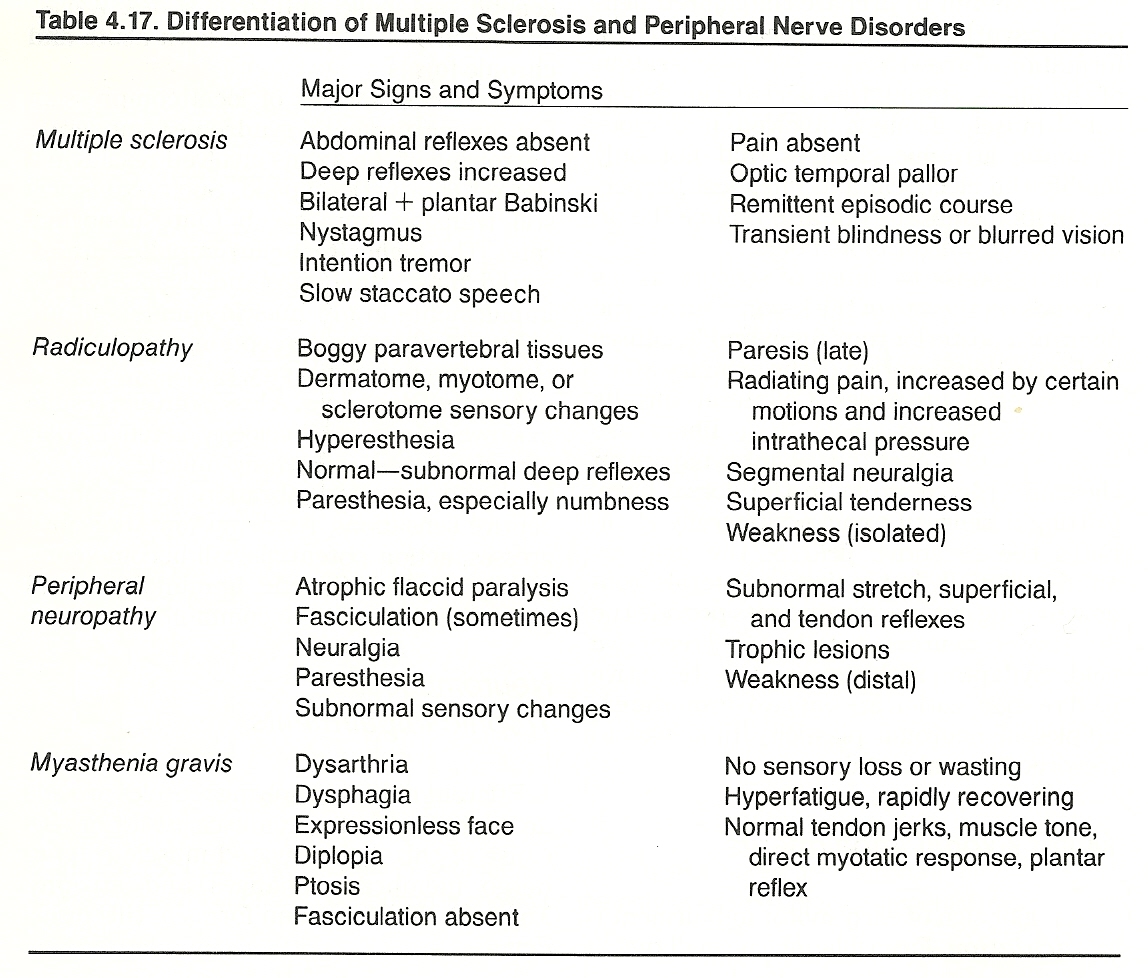
Peripheral Nerve Entrapment Syndromes
A peripheral entrapment syndrome represents a distinct type of neuropathy in which a single nerve is compressed at a specific site (eg, within fibrous tissue, a fibrous-osseous tunnel, or a muscle), either by external forces or by normally surrounding tissues. Local impairment of blood supply may further damage the entrapped nerve if associated vessels become stretched, kinked, or compressed. Thus, two factors must be considered:
(1) the effects of pressure upon nerve fibers and
(2) the effects of ischemia upon nerve fibers. These factors may exist separately or in combination.
So that a patient may avoid unnecessary pain and disability, it is important to identify a peripheral entrapment syndrome rapidly through careful examination and appropriate diagnostic studies such as EMGs, nerve conduction evaluations, and diagnostic imaging. Severe impairment of nerve function is usually only reversible in its early stages.
The major features of upper- and lower-extremity nerve entrapment syndromes are shown in Table 4.18.
Spinal entrapment syndromes are described throughout Part III. (Chapters 5-10)